Research
Introduction
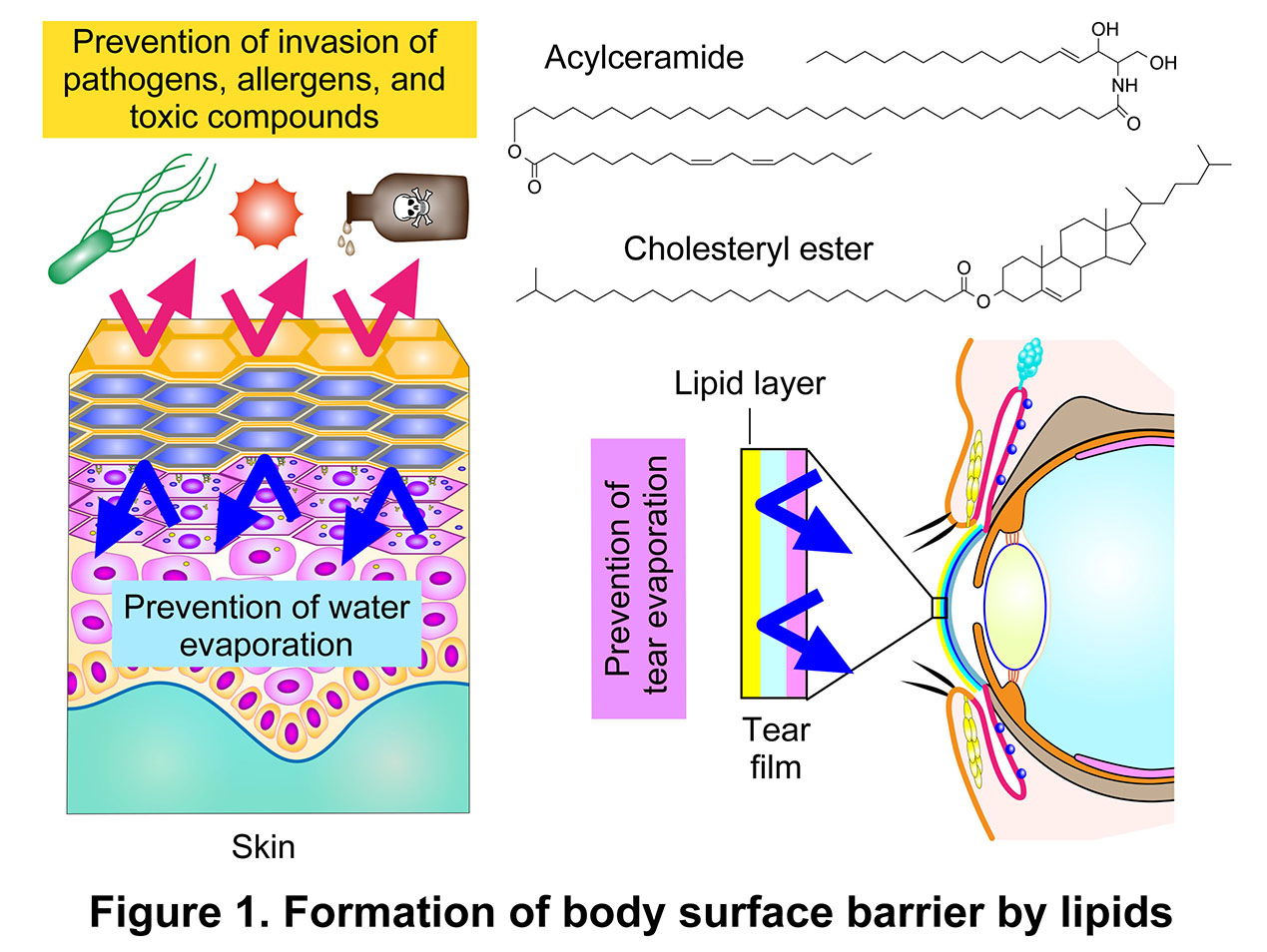
The Laboratory of Biochemistry focuses on lipids. Lipids are defined as water insoluble biomolecules, and living organisms contain a wide variety of lipid classes and species. Beyond the three major functions traditionally attributed to lipids—energy storage, membrane formation, and lipid mediator function—our laboratory focuses on the barrier function, which we propose as an additional major function of lipids (Figure 1). At body surfaces such as the epidermis and the tear film, lipids form permeability barriers that restrict the penetration of pathogens, allergens, and other external substances and prevent excessive water loss (Figure 2). Barrier function is mediated by specialized lipid classes that are structurally optimized for this purpose. In the epidermis, unique ceramides, including acylceramides, are essential for skin barrier formation, whereas in the tear film lipid layer, barrier function is primarily mediated by meibum lipids, including cholesteryl esters. These barrier-forming lipids share exceptionally long carbon chains, conferring strong hydrophobicity that enables the formation of highly impermeable lipid layers. Impairment of lipid-mediated barrier function is closely associated with human diseases. In the skin, impairment of the barrier increases the risk of infections and contributes to the development of atopic dermatitis and ichthyosis (currently referred to as epidermal differentiation disorders; EDD). In the eye, dysfunction of the tear film lipid barrier is associated with dry eye disease and, in severe cases, corneal disorders. For detailed descriptions of skin and tear film barriers, please refer to Section 4: Ceramide-mediated skin barrier formation and Section 5: Prevention of dry eye by meibum lipids . The Laboratory of Biochemistry also investigates the physiological and pathological roles and metabolism of various other lipids, including complex sphingolipids as well as ether lipids such as plasmalogens and seminolipids.
Research Overview
Lipids exhibit far greater diversity in structure than other biomolecules such as amino acids, nucleic acids, and sugars. In mammals, lipids comprise three major classes: glycerolipids, sphingolipids, and sterols. Even differences in polar head groups alone further subdivide these classes, and among them, sphingolipids are particularly diverse and include sphingomyelin and hundreds of glycosphingolipids with distinct sugar chain structures (Figure 3). Lipid diversity is not limited to polar head groups. Variations in hydrophobic moieties, including chain length, degree of unsaturation, and the presence or absence of hydroxyl groups, expand the number of lipid species to an estimated several hundred thousand. This extraordinary diversity has expanded over the course of biological evolution. In lower organisms such as bacteria, sterols and sphingolipids are mostly absent, with only a few exceptions. If lipids functioned only to compartmentalize cells and organelles by forming membranes, a relatively small number of lipid species would be sufficient, as seen in bacteria. However, as organisms diversified over long evolutionary timescales, with some lineages evolving multicellularity and adapting to complex environments, a wide variety of organisms, tissues, and cell types emerged. In parallel, lipids are thought to have diversified into an enormous number of molecular species specialized for particular organisms, tissues, and cell types.
The Laboratory of Biochemistry conducts research on diverse lipids. Our work originally centered on sphingolipids, although our interests have since broadened to encompass other lipid types, including meibum lipids. The hydrophobic backbone of sphingolipids is ceramide, which consists of a long-chain base amide-linked to a fatty acid (Figure 3). Sphingolipids play essential roles in a wide range of physiological processes, including skin barrier formation, neural function, glucose homeostasis, pathogen recognition, immunity, angiogenesis, bone formation, and spermatogenesis. Consequently, dysregulation of sphingolipid metabolism is associated with diverse pathological conditions, such as skin disorders, neurological diseases, metabolic syndrome, and cancer.

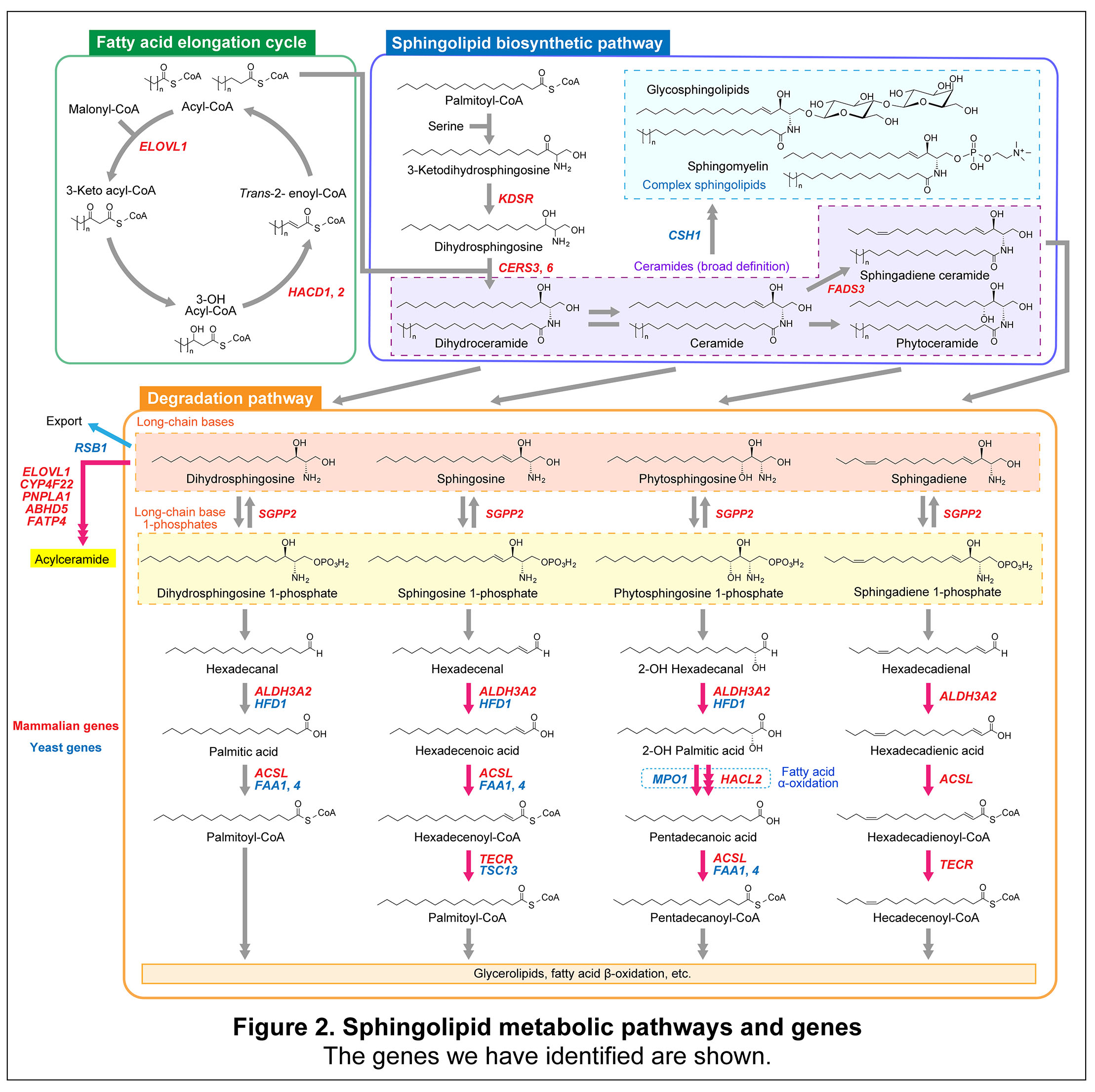
To date, the Laboratory of Biochemistry has identified and characterized numerous genes involved in the biosynthesis, modification, and degradation of sphingolipids (see Section 1: Identification of sphingolipid-related genes, for details). With regard to degradation, our laboratory has focused on the metabolic pathways of long-chain bases, which are the core structural components of sphingolipids. Within these pathways, we identified all genes encoding enzymes acting downstream of sphingosine 1-phosphate (S1P) lyase, starting from the fatty aldehyde dehydrogenase step, thereby completing our understanding of long-chain base metabolism beyond S1P cleavage (see Section 2: Metabolic pathways of long-chain bases and fatty acid α-oxidation, for details).
S1P is a well-known lipid mediator present in extracellular fluids, such as plasma and lymph, and plays critical roles in the vascular and immune systems. Fingolimod (development code FTY720), an immunomodulatory drug that exploits the immunological functions of S1P, has already been approved for the treatment of multiple sclerosis. In addition to its role as a lipid mediator in mammals, long-chain base phosphates, including S1P, serve as key metabolic intermediates in long-chain base metabolism. This metabolic role is conserved from yeast to humans and is evolutionarily more ancient and universal than the function of S1P as a lipid mediator.
In mammals, the major long-chain base is sphingosine, which serves as the precursor of S1P. Another long-chain base, phytosphingosine, which possesses a hydroxyl group at the C4 position, is present in specific tissues such as the skin, small intestine, and kidney. The Laboratory of Biochemistry has elucidated the metabolic pathway of phytosphingosine, revealing an α-oxidation pathway in which phytosphingosine is converted into odd-chain fatty acids via 2-hydroxy fatty acid intermediates; for example, C18 phytosphingosine is metabolized into pentadecanoic acid. Furthermore, our laboratory has elucidated the biosynthetic pathway of the long-chain base 4,14-sphingadiene and identified FADS3 as the gene encoding the desaturase that introduces a cis double bond at the C14 position of sphingosine-containing ceramides.
Fatty acids are classified as long-chain fatty acids (LCFAs; C11–20) and very-long-chain fatty acids (VLCFAs; ≥C21). Glycerolipids predominantly contain LCFAs, most commonly C16, C18, and C20 species. In contrast, sphingolipids are characterized by a high prevalence of VLCFAs, particularly C22 and C24 species, although they also contain abundant C16 and C18 LCFAs. VLCFAs are synthesized through the fatty acid elongation cycle in the endoplasmic reticulum. The Laboratory of Biochemistry investigates the molecular mechanisms of the fatty acid elongation cycle, as well as the physiological functions of lipids containing VLCFAs and their associations with diseases (see Section 3: Production, functions, and pathology of very-long-chain fatty acids, for details).
Ceramides are present throughout the body; however, because they function as metabolic intermediates in sphingolipid biosynthesis, their abundance is generally limited in most tissues. An important exception is the epidermis, where ceramides are present in large quantities and exhibit remarkable structural diversity. The Laboratory of Biochemistry has established a comprehensive LC-MS/MS–based analytical method that enables quantitative and qualitative profiling of ceramides. Using this approach, we demonstrated that the human stratum corneum contains 23 ceramide classes comprising a total of 1,581 distinct species. Epidermal ceramides are enriched in species containing ultra-long-chain fatty acids (≥C26; ULCFAs). In addition, a unique class of ceramides known as acylceramides contains ULCFAs of C30–C36 whose ω-termini are hydroxylated and esterified with linoleic acid. Ceramides, and acylceramides in particular, are critical lipids for skin barrier function. Quantitative and qualitative alterations in epidermal ceramides and acylceramides lead to impairment of the skin barrier and are associated with skin disorders such as ichthyosis (EDDs) and atopic dermatitis. Although the molecular mechanisms underlying acylceramide biosynthesis had long remained incompletely understood, the Laboratory of Biochemistry has identified five of the seven genes involved in acylceramide production (see Section 4: Ceramide-mediated skin barrier formation, for details).
In the tear film, barrier function is primarily conferred by meibum lipids secreted by the meibomian glands within the tarsal plate of the eyelids. This lipid barrier is essential for maintaining corneal health and preventing dry eye disease. The tear film consists of an outer lipid layer and an inner muco-aqueous layer. The lipid layer is composed predominantly of meibum lipids, with minor contributions from phospholipids derived from other tissues, such as the corneal epithelium and lacrimal glands. The quantitatively dominant lipid classes in meibum lipids are cholesteryl esters and wax monoesters with low polarity, while more polar lipid species, including wax diesters, O-acyl-ω-hydroxy fatty acids (OAHFAs), and cholesteryl OAHFAs, are also present. It is proposed that these meibum lipids, together with phospholipids, are spatially organized from the muco-aqueous layer toward the lipid layer in order of increasing hydrophobicity, thereby forming a stable interface between the lipid and mucin layers. The Laboratory of Biochemistry investigates the molecular mechanisms underlying meibum lipid production and their roles in preventing dry eye disease (see Section 5: Prevention of dry eye by meibum lipids, for details).
The meibomian glands that produce meibum lipids are a specialized type of sebaceous gland and share anatomical and functional similarities with cutaneous sebaceous glands. Accordingly, sebum lipids partially overlap with meibum lipids in their composition. Sebum plays a particularly important role in skin barrier formation in fur-covered mammals. In addition to its central role in skin barrier function, including water repellency and antimicrobial defense, sebum contributes to other physiological processes such as thermoregulation and hair maintenance. Alterations in the amount or composition of sebum lipids are associated with various skin disorders, such as acne vulgaris, dry skin, seborrheic dermatitis, and alopecia. The Laboratory of Biochemistry investigates the diversity of sebum lipids and their roles in skin barrier function and hair maintenance (see Section 6: Barrier formation and hair maintenance by diverse sebum lipids, for details).
Wax esters present in both meibum lipids and sebum lipids contain fatty alcohol moieties. Through analyses of knockout (KO) mice lacking the genes encoding the acyl-CoA reductases FAR1 and FAR2, which are responsible for fatty alcohol biosynthesis, the Laboratory of Biochemistry has expanded its research to other classes of fatty alcohol-containing lipids, namely ether lipids. These include plasmalogens, which are widely distributed across diverse tissues, and seminolipids, which play essential roles in spermatogenesis (see Section 7: Physiological functions and biosynthesis of ether lipids, for details).
Review Articles by Our Laboratory
- Hirabayashi T, Murakami M, Kihara A. (2019) The role of PNPLA1 in ω-O-acylceramide synthesis and skin barrier function. Biochim Biophys Acta Mol Cell Biol Lipids, 1864, 869-879.
- Kihara A (2016) Synthesis and degradation pathways, functions, and pathology of ceramides and epidermal acylceramides. Prog Lipid Res, 63, 50-69.
- Kihara A (2015) Sphingolipid metabolism via sphingosine 1-phosphate and its role in physiology pathology, and nutrition. "Bioactive lipid mediators: current reviews and protocols (Yokomizo T, Murakami M eds.)", Springer Japan, Tokyo. pp 127-138.
- Kihara A (2014) Sphingosine 1-phosphate is a key metabolite linking sphingolipids to glycerophospholipids. Biochim Biophys Acta, 1841, 766-772.
- Sassa T, Kihara A (2014) Metabolism of very long-chain fatty acids: genes and pathophysiology. Biomol Ther, 22, 83-92.
- Kihara A (2012) Very long-chain fatty acids: elongation, physiology and related disorders. J Biochem, 152, 387-395.
- Kihara A, Igarashi Y (2008) Production and release of sphingosine 1-phosphate and the phosphorylated form of the immunomodulator FTY720. Biochim Biophys Acta, 1781, 496-502.
- Kihara A, Mitsutake S, Mizutani Y, Igarashi Y (2007) Metabolism and biological functions of two phosphorylated sphingolipids, sphingosine 1-phosphate and ceramide 1-phosphate. Prog Lipid Res, 46, 126-144.