Research
研究内容
3. 極長鎖脂肪酸の産生,生理機能,病態
極長鎖脂肪酸の定義は必ずしも統一されていないが,ここでは最も一般的な定義である C(炭素数)21 以上とする。また,極長鎖脂肪酸のうち,C26 以上のものはそれより短い極長鎖脂肪酸と機能的に異なっているので超長鎖脂肪酸と呼んで区別することがあり,ここでもそのように記載する。極長鎖脂肪酸(超長鎖脂肪酸を含む)の細胞内含量は全体の脂肪酸量から見ると数パーセントにしか満たないが,長鎖脂肪酸(C11−20)では代替できない重要な生理機能を有する1-4)。例えば,皮膚には C30 を超える超長鎖脂肪酸を含むセラミド(アシルセラミドなど)が存在し,皮膚バリア機能に必須である1-5)。C24 極長鎖脂肪酸を含有するスフィンゴ脂質(C24 スフィンゴ脂質)は広く哺乳類組織に分布しているが,特に肝臓やミエリンに多く存在する。C24 スフィンゴ脂質を産生できない KO マウス(セラミド合成酵素 Cers2 KO マウス)では肝臓の機能異常・肝癌発症,神経での脱髄が見られることが報告されている。また,C24 スフィンゴ脂質はマイクロドメイン機能に重要である。C28 を超える多価不飽和超長鎖脂肪酸を含有したスフィンゴ脂質は精子特異的に存在し,精子の成熟や受精能獲得において重要である。脳や網膜ではホスファチジルコリン(PC)の sn-2 位にドコサヘキサエン酸(C22:6)が多く見られる。また,これらの臓器のホスファチジルコリンの sn-1 位には他の臓器では見られない C28 以上の多価不飽和超長鎖脂肪酸が存在する。超長鎖脂肪酸含有ホスファチジルコリンは神経の発達や機能,網膜機能に重要な働きを示す。ドコサヘキサエン酸はグリセロリン脂質の構成成分としてだけではなく,プロテクチンやレゾルビンといった抗炎症・炎症収束に働く脂質メディエーターの前駆体としても機能する。
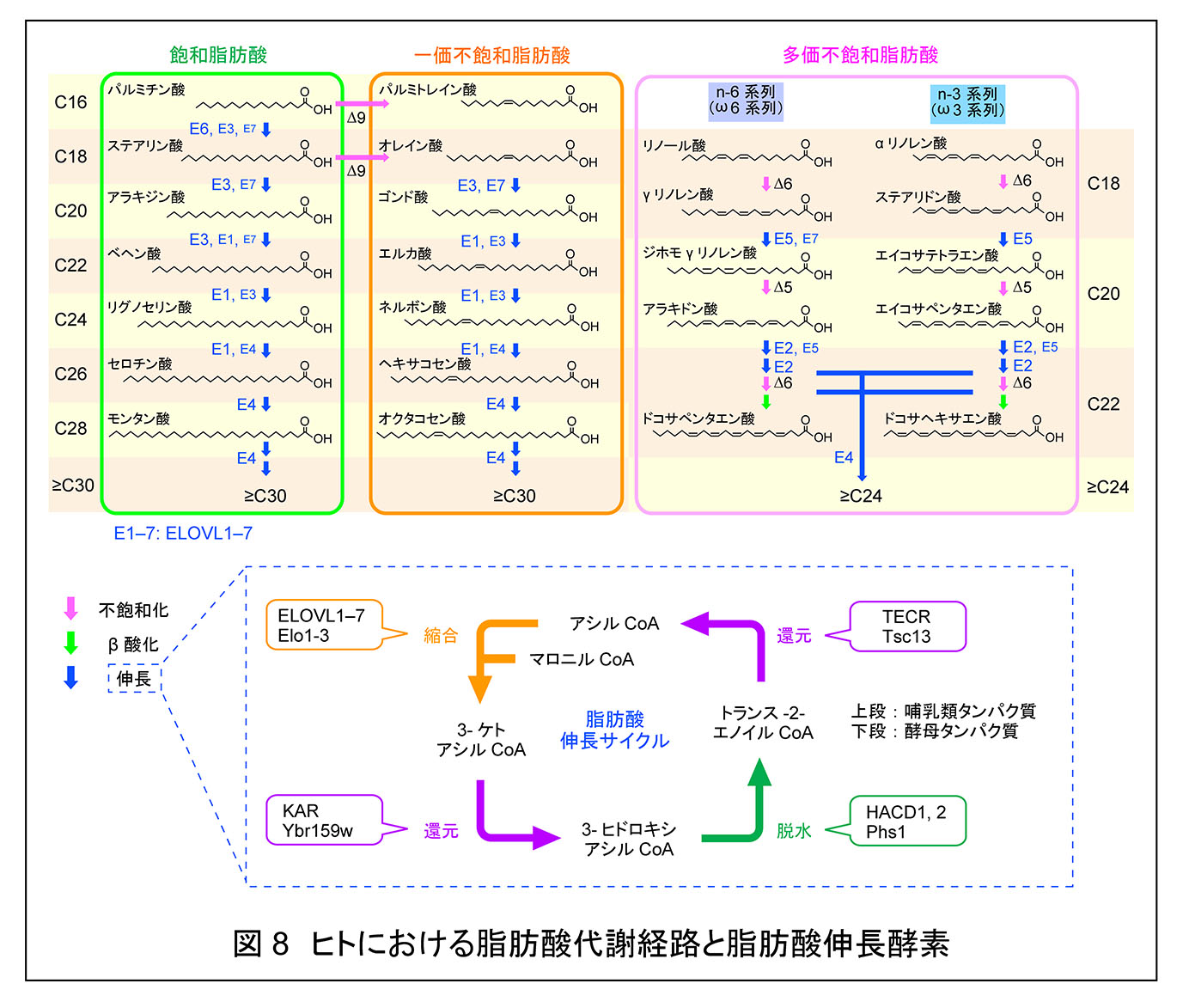
脂肪酸の伸長は小胞体で行われる。脂肪酸はアシル CoA に変換後,縮合,還元,脱水,還元の4つのステップを 1 サイクルとした脂肪酸伸長サイクルによって炭素数 2 ずつ伸長される(図 8)。最初の縮合ステップが律速段階であり,脂肪酸伸長酵素(エロンガーゼ)によって触媒される。哺乳類には 7 種の脂肪酸伸長酵素(ELOVL1−7)が存在し,それぞれが異なった基質特異性を示す。生化学研究室は 様々なアシル CoA 基質を用いた in vitro での解析によって,各 ELOVL の基質特異性を明らかにした6, 17)(図 8)。また,この実験によって ELOVL1 が C24 スフィンゴ脂質産生に中心的な役割を果たすことが明らかとなった。ELOVL1 をノックダウンした HeLa 細胞 では,C24 スフィンゴ脂質の量が減少し,C16 スフィンゴ脂質量が増加した。このような細胞では Src ファミリー Lyn の活性化が抑制され6),アポトーシスが亢進する7)など細胞のシグナル伝達に変化が生じていた。また,生化学研究室では in vivo での Elovl1 の生理的役割を解析するために Elovl1 KO マウスを作成し,皮膚バリア不全のため新生致死となることを明らかにした8)。KO マウスでは表皮における C26 以上の超長鎖(非アシル化)セラミドとアシルセラミド,脳や肝臓などの表皮以外の組織における C22,C24 極長鎖スフィンゴ脂質の減少が観察された。ほとんどの組織におけるスフィンゴ脂質の脂肪酸部分の多くは飽和脂肪酸であるが,例外的に C24 には C24:0 の他に C24:1 と C24:2 が比較的多く存在し,これらの産生にも ELOVL1 が働くことを見出した9)。これらの不飽和スフィンゴ脂質は飽和タイプとは異なり,脂質マイクロドメインの外に局在する傾向が強い。
極長鎖脂肪酸の β 酸化はペルオキシソームで行われる。極長鎖アシル CoA のペルオキシソームの輸送には ABC トランスポーター ABCD1 が関与する。X 染色体に存在するこの遺伝子の変異は極長鎖脂肪酸の蓄積を引き起こし,副腎白質ジストロフィー(X-ALD)を引き起こす。ロレンツォオイルはオレイン酸(C18:1)とエルカ酸(C22:1)からなるトリグリセリドの 4:1 混合物であり,血漿中の飽和極長鎖脂肪酸(C24:0, C26:0)を減少させる効果がある。これまでロレンツォオイルがどのようなメカニズムで極長鎖脂肪酸の蓄積を抑えるのかは不明であったが,生化学研究室では ELOVL1 の活性阻害を介して飽和極長鎖脂肪酸の産生を低下させることを明らかにした10)。ロレンツォオイルは血漿中の飽和極長鎖脂肪酸を減少させる効果があるが,不飽和極長鎖脂肪酸の蓄積の軽減や治療薬としての効果については問題が残る。我々の結果は ELOVL1 が X-ALD 治療薬のターゲットとしては極めて有用であると示しているので,今後よりよい ELOVL1 阻害剤の開発が待たれる。
ELOVL4 は炭素数 28 以上の超長鎖脂肪酸の産生に関わる。これらの脂質は生体において,皮膚(アシルセラミドなど),脳(PC),網膜(PC),精子(セラミド,スフィンゴミエリン),マイボーム腺(コレステリルエステル,ワックスエステルなどのマイバム脂質)に存在する。ELOVL4 遺伝子の変異はこれらの組織における機能異常を引き起こし,黄斑変性疾患,神経疾患,皮膚魚鱗癬の発症につながる。ELOVL4 遺伝子変異に起因する疾患はELOVL4 変異は変異の場所や種類により,3つのタイプ(スタルガルト病 3 型,脊髄小脳性運動失調症(SCA)34 型,ISQMR)に分類される。このうち,SCA34 は小脳の萎縮と運動失調を特徴とする常染色体顕性遺伝性疾患である。生化学研究室では SCA34 の顕性遺伝性様式が超長鎖 PC の鎖長の低下によって引き起こされることを見出した23)。
ELOVL7 は C18−20 のアシル CoA に対して活性を示す。生化学研究室では精製した ELOVL7 をプロテオリポソームに再構成することで,哺乳類脂肪酸伸長酵素としては初めて縮合活性を測定することに成功した11)。脂肪酸伸長サイクルの第一段階目は ELOVL が触媒するが,第二段階目の還元反応は KAR が触媒する。生化学研究室では C16:0-CoA に活性を示す ELOVL6 と KAR のプロテオリポソームへの再構成により,KAR の存在下で ELOVL が高い活性を示すことを示した。このことは脂肪酸伸長サイクルの第一段階と第二段階が協調して働いていることを示している12)。
生化学研究室では脂肪酸伸長サイクルの第三段階の反応に関わる 3-ヒドロキシアシル CoA 脱水素酵素として HACD1−4 を同定した13)。これらはいずれも in vitro の実験において3-ヒドロキシパルミトイル CoA に対して活性を示した。一方,HACD をノックアウトした細胞を用いた実験から,HACD1 と HACD2 が幅広い 3-ヒドロキシアシル CoA 基質に対して活性を示すこと,HACD2 がほとんどの組織で主要な 3-ヒドロキシアシル CoA 脱水素酵素であることを明らかにした14)。HACD3 と HACD4 の活性は細胞レベルでは弱いか,ほとんど検出されなかった。HACD1 は心臓や骨格筋など,筋肉系組織特異的に発現しており,イスラエルとの共同研究によってヒトミオパチーの原因遺伝子であることを明らかにした15)。脂肪酸伸長サイクルの第四段階を触媒する TECR の P182L 変異は非症候性精神遅滞の原因遺伝子であることが知られている。生化学研究室ではこの変異がタンパク質の安定性と活性を低下させ,細胞内の C24 極長鎖スフィンゴ脂質量を低下させることを明らかにした16)。また,これらの解析を通じて TECR の活性低下が脂肪酸伸長サイクル第三段階目を触媒する HACD の活性に影響を与えることを見いだした。このことは肪酸伸長サイクルにおいて第三段階と第四段階が協調して働いていることを示している。TECR P182L 変異タンパク質は活性を完全には失っておらず,脂肪酸伸長サイクルに与える影響という点では比較的弱い。このような弱い変異によって神経系が影響を受けるということは,神経系の極長鎖脂肪酸の依存度が極めて高いということを示唆している。神経細胞の軸索を覆うミエリンには脂質が豊富に存在し,絶縁性を高めている。脂質の中でも特に C24 の鎖長をもつガラクトシルセラミド,スルファチドなどの極長鎖スフィンゴ脂質はミエリンの形成・機能維持に重要であり,TECR P182L 変異はこれらの C24 スフィンゴ脂質量の低下を介してミエリン機能に異常を与えていると予測される。長い脂肪酸鎖長は曲率の高い膜構造体を安定化するのであろう(後述)。トランス-2-エノイル CoA 還元酵素(酵母 Tsc13,哺乳類 TECR)の触媒基や反応機構は不明であったが,生化学研究室では Tsc13/TEC Rの Tyr256/Tyr248 が触媒基として働き,トランス-2-エノイル CoA へのプロトンを供与していることを明らかにした24)。さらに,Tsc13 と脂肪酸伸長サイクルの第一段階を触媒する縮合酵素(Elo2/3)との相互作用について明らかとし,その相互作用が縮合酵素の活性発現に重要であることを解明した。すなわち,脂肪酸伸長サイクルにおいても上述の第一段階と第三段階および第三段階と第四段階だけでなく,第四段階と引き続く第一段階も協調して働いている。
上述のように全身の Elovl1 遺伝子を欠損したマウスは皮膚バリア形成不全によって新生致死であったため,表皮以外の組織における極長鎖脂肪酸の役割の解明には至らなかった。そこで,生化学研究室では表皮で Elovl1 遺伝子が発現するものの,表皮以外の組織では Elovl1 遺伝子が欠損したマウスを作成し,解析を行った。その結果,このマウスの目は若齢期においてドライアイになり,5ヶ月齢以降では角膜が混濁した17)。涙液バリアの詳細については「5. マイバム脂質によるドライアイ防止」の項を参照されたい。また,ヒトにおいて ELOVL1 変異が皮膚魚鱗癬,下肢痙性,歩行困難,ミエリン低形成などを伴う神経皮膚疾患を引き起こすことを明らかにした18, 25)。Elovl1 変異マウスにおいても運動障害,ミエリン低形成が観察された19)。この変異マウスの脳ミエリンではミエリンの機能維持に重要なガラクトシルセラミドの量の低下と各種スフィンゴ脂質(ガラクトシルセラミド,スルファチド,スフィンゴミエリン)の短鎖化が見られ,これらが神経症状の原因となっていると考えられる。
極長鎖脂肪酸は酵母からヒトに至るまで真核生物に広く保存されている。酵母 Saccharomyces cerevisiae の極長鎖脂肪酸はほぼ全てが C26 であり,そのほとんどがスフィンゴ脂質合成に使用される。極長鎖脂肪酸合成が不全となった酵母は致死となることから,極長鎖脂肪酸が生育に必須な役割を果たすことは明らかである。脂肪酸伸長サイクルの 4 つの反応を触媒する酵素はいずれも真核生物で保存されており,酵母 Phs1 は哺乳類 HACD1−4 のホモログである。生化学研究室では Phs1 の活性に重要な残基を同定し,反応機構に関する知見を得た20, 21)。また,極長鎖脂肪酸の分子レベルでの機能を明らかにするため,極長鎖脂肪酸が低下して高温で生育ができなくなった酵母を用いた遺伝学的な解析を行った。その結果,この条件下での生育不全をレスキューするマルチコピーサプレッサーとして後期エンドソーム(マルチベシキュラーボディー;MVB)を介した小胞輸送に関わる VPS21 を単離した22)。この結果は,極長鎖脂肪酸(極長鎖脂肪酸含有スフィンゴ脂質)が MVB を介した小胞輸送において重要な機能を果たすことを示唆している。極長鎖脂肪酸はその長さから脂質二重層の両層にまたがって局在することにより,曲率の高い不安定な膜構造の安定化に働くと考えられる。MVB では内腔小胞が形成される際にこのような曲率の高い膜構造体が多く作られるので,極長鎖脂肪酸の必要性が高いのであろう。また,上述のように,TECR やELOVL1 の変異は神経症状を引き起こす。ミエリンは軸索の周りに脂質の層が何重にも巻き付いた曲率の高い構造をしており,この安定化に極長鎖型のスフィンゴ脂質が寄与していると考えられる。
研究業績
- 木原章雄 (2018) 鎖長の違いによる脂肪酸クオリティ — 極長鎖・超長鎖脂肪酸の産生機構と病態. 医学のあゆみ, 264, 966-971. Review
- Kihara A (2016) Synthesis and degradation pathways, functions, and pathology of ceramides and epidermal acylceramides. Prog Lipid Res, 63, 50-69. Review.
- Sassa T, Kihara A (2014) Metabolism of very long-chain fatty acids: genes and pathophysiology. Biomol Ther, 22, 83-92. Review.
- Kihara A (2012) Very long-chain fatty acids: elongation, physiology and related disorders. J Biochem, 152, 387-395. Review
- 木原章雄 (2015) セラミドによる皮膚バリア形成. 実験医学, 33, 117-122. Review.
- Ohno Y, Suto S, Yamanaka M, Mizutani Y, Mitsutake S, Igarashi Y, Sassa T, Kihara A (2010) ELOVL1 production of C24 acyl-CoAs is linked to C24 sphingolipid synthesis. Proc Natl Acad Sci USA, 107, 18439-18444.
- Sassa T, Suto S, Okayasu Y, Kihara A (2012) A shift in sphingolipid composition from C24 to C16 increases susceptibility to apoptosis in HeLa cells. Biochim Biophys Acta, 1821, 1031-1037.
- Sassa T, Ohno Y, Suzuki S, Nomura T, Nishioka C, Kashiwagi T, Hirayama T, Akiyama M, Taguchi R, Shimizu H, Itohara S, Kihara A (2013) Impaired epidermal permeability barrier in mice lacking the Elovl1 gene responsible for very long-chain fatty acid production. Mol Cell Biol, 33, 2787-2796.
- Edagawa M, Sawai M, Ohno Y, Kihara A (2018) Widespread tissue distribution and synthetic pathway of polyunsaturated C24:2 sphingolipids in mammals. Biochim Biophys Acta, 1863, 1441-1448.
- Sassa T, Wakashima T, Ohno Y, Kihara A (2014) Lorenzo's oil inhibits ELOVL1 and lowers the level of sphingomyelin with a saturated very long-chain fatty acid. J Lipid Res, 55, 524-530.
- Naganuma T, Sato Y, Sassa T, Ohno Y, Kihara A (2011) Biochemical characterization of the very long-chain fatty acid elongase ELOVL7. FEBS Lett, 585, 3337-3341.
- Naganuma T, Kihara A (2014) Two modes of regulation of the fatty acid elongase ELOVL6 by the 3-ketoacyl-CoA reductase KAR in the fatty acid elongation cycle. PLoS One, 9, e101823.
- Ikeda M, Kanao Y, Yamanaka M, Sakuraba H, Mizutani Y, Igarashi Y, Kihara A (2008) Characterization of four mammalian 3-hydroxyacyl-CoA dehydratases involved in very long-chain fatty acid synthesis. FEBS Lett, 582, 2435-2440.
- Sawai M, Uchida Y, Ohno Y, Miyamoto M, Nishioka C, Itohara S, Sassa T, Kihara A (2017) The 3-hydroxyacyl-CoA dehydratases HACD1 and HACD2 exhibit functional redundancy and are active in a wide range of fatty acid elongation pathways. J Biol Chem, 292, 15538-15551.
- Muhammad E, Reish O, Ohno Y, Scheetz T, Deluca A, Searby C, Regev M, Benyamini L, Fellig Y, Kihara A, Sheffield VC, Parvari R (2013) Congenital myopathy is caused by mutation of HACD1. Hum Mol Genet, 22, 5229-5236.
- Abe K, Ohno Y, Sassa T, Taguchi R, Çalış kan M, Ober C, Kihara A (2013) Mutation for nonsyndromic mental retardation in the trans-2-enoyl-CoA reductase TER gene involved in fatty acid elongation impairs the enzyme activity and stability, leading to change in sphingolipid profile. J Biol Chem, 288, 36741-36749.
- Sassa T, Tadaki M, Kiyonari H, Kihara A (2018) Very long-chain tear film lipids produced by fatty acid elongase ELOVL1 prevent dry eye disease in mice. FASEB J, 32, 2966-2978.
- Mueller N, Sassa T, Morales-Gonzalez S, Schneider J, Salchow DJ, Seelow D, Knierim E, Stenzel W, Kihara A, Schuelke M (2019) De novo mutation in ELOVL1 causes ichthyosis, acanthosis nigricans, hypomyelination, spastic paraplegia, high frequency deafness and optic atrophy. J Med Genet, 56, 164-175.
- Isokawa M, Sassa T, Hattori S, Miyakawa T, Kihara A (2019) Reduced chain length in myelin sphingolipids and poorer motor coordination in mice deficient in the fatty acid elongase Elovl1. FASEB Bioadv, 1, 747-759.
- Kihara A, Sakuraba H, Ikeda M, Denpoh A, Igarashi Y (2008) Membrane topology and essential amino acid residues of Phs1, a 3-hydoroxyacyl-CoA dehydratase involved in very long-chain fatty acid elongation. J Biol Chem, 283, 11199-11209.
- Yazawa T, Naganuma T, Yamagata M, Kihara A (2013) Identification of residues important for the catalysis, structure maintenance, and substrate specificity of yeast 3-hydroxyacyl-CoA dehydratase Phs1. FEBS Lett, 587, 804-809.
- Obara K, Kojima R, Kihara A (2013) Effects on vesicular transport pathways at the late endosome in cells with limited very long-chain fatty acids. J Lipid Res, 54, 831-842.
- Tamura Y, Sassa T, Nishizawa T, Kihara A. (2023) Incomplete elongation of ultra-long-chain polyunsaturated acyl-CoAs by the fatty acid elongase ELOVL4 in spinocerebellar ataxia type 34. Mol Cell Biol, 43, 85-101.
- Kato R, Takenaka Y, Ohno Y, Kihara A. (2024) Catalytic mechanism of trans-2-enoyl-CoA reductases in the fatty acid elongation cycle and its cooperative action with fatty acid elongases. J Biol Chem, 300, 105656.
- Takahashi T, Turker S, Sassa T, Akcapinar GB, Yararbas K, Susguna S, Iseri SAU, Kihara A, Akcakaya NH. (2022) Hypomyelinating spastic dyskinesia and ichthyosis caused by a homozygous splice site mutation leading to exon skipping in ELOVL1. Brain Dev, 44, 391-400.