Research
研究内容
5. マイバム脂質によるドライアイ防止
涙液は油層と粘液層(水層,ムチン層)から構成されており(図 13),最も外側にある油層は水分蒸発の防止や表面張力の低下という役割をもつ。涙液油層に存在する脂質は瞼の裏側にあるマイボーム腺から分泌されるため,総称してマイバム脂質と呼ばれる。近年,パソコンやスマートフォンの普及,エアコンの使用などの生活環境の変化に伴い,ドライアイ患者が増加している。ドライアイの約8割はマイボーム腺の機能異常,すなわち油層の異常であることが知られているが,現在のドライアイの治療薬には水層とムチン層をターゲットにしたものしか存在しない。
マイバム脂質の主成分はコレステリルエステル(30−67%)とワックスモノエステル(30−48%)であり,共に炭素鎖長(コレステリルエステルのアルコール部分,ワックスモノエステルの脂肪酸部分)が長い(C20−34)という特徴をもつ。これらの脂質の極性は極めて低く,油層の中でも非極性サブレーヤーを構成すると考えられている(図 13)。一方,マイバム脂質には O-アシル-ω-水酸化脂肪酸(OAHFA)などの両親媒性の脂質も少ないながら(1−5%)存在し,非極性サブレーヤーと水層の間に両親媒性サブレーヤーを形成し,両層を繋ぎ止める役割があると推測されている。しかし,これまで涙液油層に関する研究は極めて少なく,マイバム脂質産生の分子機構,個々の脂質の役割,性質上馴染むことができない水と脂質が涙液で安定的に維持されているメカニズムなど,不明な点が多く残されていた。

生化学研究室では,極長鎖脂肪酸の産生に働く脂肪酸伸長酵素 Elovl1 の変異マウス(新生致死性を回避するため,表皮以外の組織で Elovl1 が欠損するように設計されたマウス)がドライアイを示すことを明らかにした1)(プレスリリース原稿)。このマウスは若齢期から瞬きの増加,眼球からの水分蒸散の亢進,涙液層の不安定化(涙液層破壊時間の短縮)を示したが,5ヶ月齢以降ではヒトの重症ドライアイ患者で観察されるような角膜混濁を示した1, 2)。マイバム脂質の主成分であるコレステリルエステルとワックスモノエステルを質量分析法(液体クロマトグラフィー連結タンデム質量分析法;LC-MS/MS)で解析した結果,コレステリルエステルの脂肪酸部分とワックスエステルのアルコール部分は共に炭素鎖長平均で約3短くなっていることが明らかとなった1, 3)。これらの結果はドライアイを防止するために涙液脂質の長さ,すなわち質(クオリティ)が重要であることを初めて示したものである。野生型マウスにおいて,マイバム脂質中のコレステリルエステルの脂肪酸鎖長は主に C16−27,ワックスモノエステルのアルコール鎖長は C24−29 であり,共に分岐型(イソ型あるいはアンテイソ型)が多い。これらの産生においては分岐アミノ酸から派生したイソブチリル CoA,イソバレリル CoA,α-メチルブチリル CoA が脂肪酸合成酵素によって長鎖にまで伸長された後に,脂肪酸伸長酵素 ELOVL3 と ELOVL1 によって極長鎖にまで伸長されることも明らかにした3)。
OAHFA は両親媒性サブレーヤーを構成し,両側にある非極性サブレーヤーと水層を安定的に繋ぎ止めていると考えられてきたが,実験的な証拠は存在しなかった。OAHFA の役割を解明するためには,まず OAHFA の産生に関わる遺伝子を明らかにし,その遺伝子変異マウスを作成することで OAHFA をもたないモデルを確立する必要があった。OAHFA はアシルセラミドから長鎖塩基を除いた構造をもつ(ただし,ω 位にエステル結合したアシル基はアシルセラミドでは主にリノール酸であり,OAHFA ではヒトでオレイン酸,マウスでパルミトオレイン酸である)。そのため,OAHFA に特徴的な ω-水酸基を導入する酵素として,アシルセラミド産生において同定した CYP4F22(マウスでは Cyp4f39)が働いていると予測された。そこで,Cyp4f39 変異マウス(新生致死性を回避するため,表皮以外の組織で Cyp4f39 が欠損するように設計されたマウス)を作成し,マイバム中での OAHFA 量を測定した結果,コントロールマウスの 1/5 まで低下したことから,CYP4F22/Cyp4f39 の OAHFA 産生への関与が示された4)(プレスリリース原稿)。Cyp4f39 変異マウスは瞬き数の亢進,涙液層破壊時間の短縮,角膜上皮障害の亢進といった典型的なドライアイ表現型を示した。
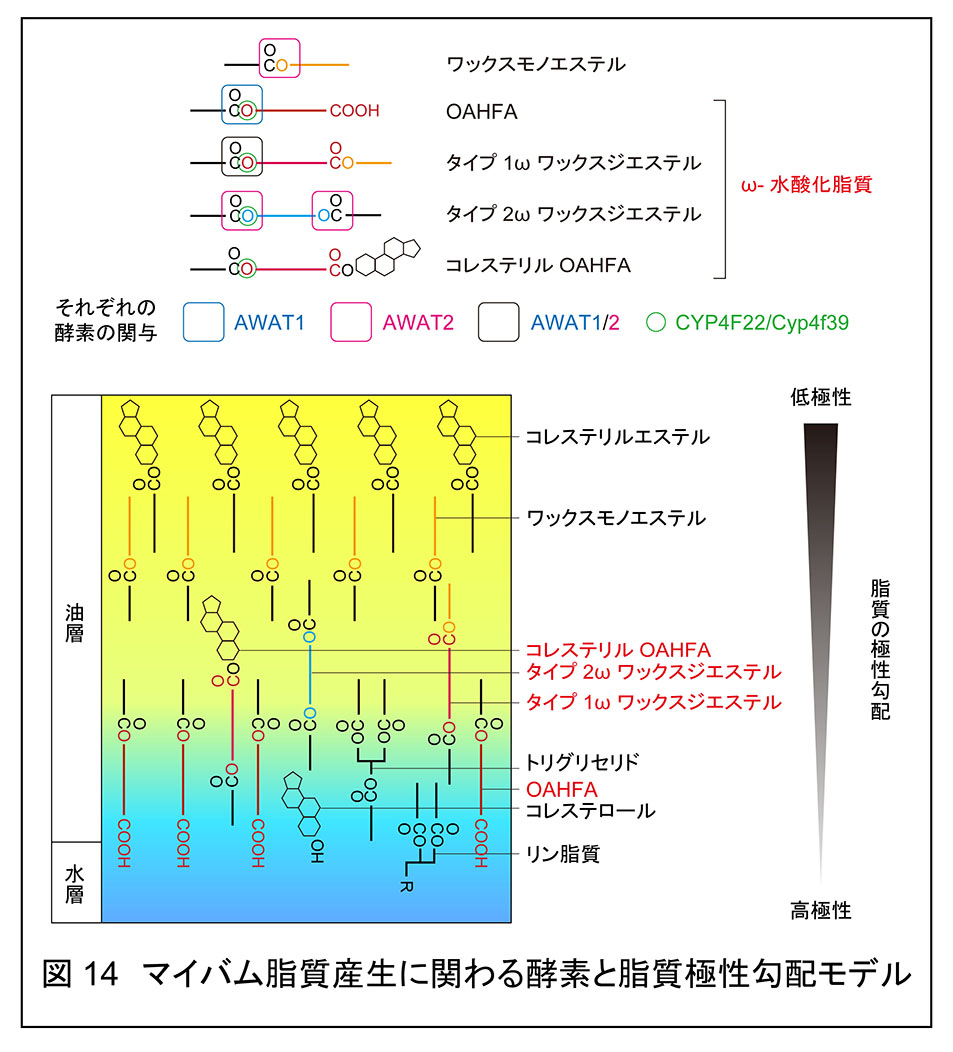
生化学研究室では,OAHFA をはじめとした多くのマイバム脂質を分離・定量する質量分析法(LC-MS/MS)を確立した。各マイバム脂質の分離・分析のためには,それぞれの脂質の標準品の存在が不可欠である。しかし,多くのマイバム脂質には標準品が存在しなかったため,生化学研究室では独自にそれらを化学合成して標準品を調製し,液体クロマトグラフィー(LC)での保持時間,タンデム質量分析におけるプロダクトイオンの設定を行った。これらにより,OAHFA の測定が可能となっただけでなく,これまで正確に測定されてこなかった他のマイバム脂質群の測定も可能となった。例えば,マイバム脂質にはワックスジエステルが存在している(約 7 %)ことが知られていたが,それらの詳細な構造は明らかになっていなかった。ワックスジエステルには,水酸化脂肪酸を骨格としたタイプ 1 とジオールを骨格としたタイプ 2 が存在し,それらはさらに水酸基の位置によって α と ω,つまりタイプ 1α あるいは ω とタイプ 2α あるいは ω に分類される。生化学研究室では,マウスマイバム脂質中にタイプ 1ω,タイプ 2α,タイプ 2ω ワックスジエステルが存在することを明らかにし,このうちタイプ 1ω,タイプ 2ω ワックスジエステルの産生が Cyp4f39 依存的であることを見出した4, 5)(図 14)。また,マイバム脂質にはコレステリル OAHFA も存在する(約 3 %)が,Cyp4f39 変異マウス中で消失していた。つまり,CYP4F22/Cyp4f39 は4つの ω-水酸化脂質(OAHFA,タイプ 1ω ワックスジエステル,タイプ 2ω ワックスジエステル,コレステリル OAHFA)の産生に関わっていることが明らかとなった。
マイバム脂質の多くはエステル結合を有す。しかし,マイバム脂質中のエステル結合の形成に関わるアシルトランスフェラーゼについては不明な点が多く残されていた。これまでに in vitro の解析から,アシル CoA ワックスアルコールトランスフェラーゼ AWAT1 および AWAT2 が長鎖のワックスモノエステルの産生活性を示すことが報告されていた。そこで,生化学研究室では AWAT1/AWAT2 のマイバム脂質産生への関与を明らかにするために,Awat1/Awat2 の単独あるいは二重 KO マウスを作成し,ドライアイ表現型とマイバム脂質組成を解析した。Awat2 の KO および二重 KO マウスはマイボーム腺の機能障害を伴う重度のドライアイ表現型を示し,Awat1 KO マウスは軽度のドライアイ表現型を示した5)(プレスリリース原稿)。これらのマウスではそれぞれ特定のマイバム脂質クラスが減少していた。すなわち,Awat1 KOマウスでは OAHFA とタイプ 1ω ワックスジエステル、Awat2 KO マウスではワックスモノエステルとタイプ 1ω および 2ω ワックスジエステル,二重 KO マウスではこれら全てがほとんど失われていた(図 14)。以上の結果から,AWAT1 と AWAT2 はそれぞれが特徴的な基質特異性を示し,これらによって多様なマイバム脂質を生成されることが明らかになった。
マイバム脂質には極長鎖の脂肪族アルコールやジオールなどのアルコール分子をもつ多彩なエステル化合物(ワックスモノエステル,ワックスジエステル)が含まれる。しかし、極長鎖アルコールやジオールに存在する 1 位水酸基の形成を担う酵素は不明であった。そこで,生化学研究室では脂肪族アルコール合成活性をもつことが知られている脂肪族アシル CoA 還元酵素 FAR1/2 に着目し,これらの酵素の極長鎖アルコール合成における活性とマイバム脂質産生における役割の解明を行った。FAR1,FAR2 過剩発現細胞を用いた解析によって,FAR1 が長鎖,FAR2 が極長アルコールの産生活性を示すことが明らかになった6)。さらに,Far2 KO マウスを作製し,このマウスがマイボーム腺の閉塞や涙液蒸散量の増加,涙液層破壊時間の短縮などの涙液蒸発亢進型ドライアイ表現型を示すことを見出した6)(プレスリリース原稿)。また,LC-MS/MS によって Far2 KO マウスのマイバム脂質を測定したところ,ワックスモノエステル,タイプ 1ω,タイプ 2α,タイプ 2ω ワックスジエステルいずれもがほぼ消失していた。以上の結果より,FAR2 がマイバム脂質中の極長鎖アルコール/ジオールの産生に関わり,ドライアイ防止に重要であることが明らかになった。
脂質は定義上,水に溶けない有機化合物の総称であり,本来,水とは馴染むことができないはずである。それにもかかわらず,涙液では水層上に油層が安定的に維持されている。この謎の解明が涙液の研究における重要な課題の一つであった。生化学研究室は,マイバム脂質の極性を比較し,OAHFA>OAHFA 以外の ω-水酸化脂質水酸化脂質>ワックスエステル,コレステリルエステルの順に極性が高いことを明らかにし,従来の水層/両親媒性サブレーヤー/非極性サブレーヤーという単純なモデルを発展させた脂質の極性勾配モデルを提唱した4)(図 14)。このモデルでは,水層側から OAHFA,OAHFA 以外の ω-水酸化脂質,ワックスエステル/コレステリルエステルという順に徐々に極性の低い脂質が配向することで脂質と水が境界面なく混ざり合う。
以上のように,生化学研究室ではこれまで不明な点の多かった,涙液油層の脂質の構造,組成,産生の分子機構,ドライアイ防止における役割について多くの新たな知見を得ることに成功し,油層形成機構,ドライアイの防止における正常な油層の形成の重要性を明らかにした。現在のドライアイ治療薬には液層をターゲットにしたものしか存在しないが,これらの知見は油層をターゲットとした新たな点眼薬の開発につながると期待される。
研究業績
- Sassa T, Tadaki M, Kiyonari H, Kihara A (2018) Very long-chain tear film lipids produced by fatty acid elongase ELOVL1 prevent dry eye disease in mice. FASEB J, 32, 2966-2978.(プレスリリース原稿)
- Watanabe K, Yoshida M, Okumura T, Sassa T, Kihara A, Uchiyama A. (2021) Improvement of evaporative dry eye with meibomian gland dysfunction in model mice by treatment with ophthalmic solution containing mineral oil. Transl Vis Sci Technol, 10, 21.
- Tanno H, Sassa T, Sawai M, Kihara A. (2021) Production of branched-chain very-long-chain fatty acids by fatty acid elongases and their tissue distribution in mammals. Biochim Biophys Acta Mol Cell Biol Lipids, 1866, 158842.
- Miyamoto M, Sassa T, Sawai M, Kihara A (2018) Lipid polarity gradient formed by ω-hydroxy lipids in tear film prevents dry eye disease. eLife, 9, e53582.(プレスリリース原稿)
- Sawai M, Watanabe K, Tanaka K, Kinoshita W, Otsuka K, Miyamoto M, Sassa T, Kihara A. (2021) Diverse meibum lipids produced by Awat1 and Awat2 are important for stabilizing tear film and protecting the ocular surface. iScience, 24, 102478.(プレスリリース原稿)
- Otsuka K, Sawai M, Kihara A. (2022) Formation of fatty alcohols—components of meibum lipids—by the fatty acyl-CoA reductase FAR2 is essential for dry eye prevention. FASEB J, 36, e22216.(プレスリリース原稿)