Research
研究内容
1. スフィンゴ脂質関連遺伝子の同定
脂質の生合成・代謝に限らず,生体内の殆ど全ての化学反応は酵素により触媒されている。 各反応を触媒する酵素の遺伝子を同定することはその後の生化学,分子生物学,遺伝学,生理学,病態解析に極めて重要である。例えば,遺伝子欠損体の作成はその遺伝子(あるいは遺伝子産物であるタンパク質/酵素)およびその酵素のプロダクト(生化学研究室の場合は脂質分子)の生理機能や関連する疾患の原因の解明につながる。また,リコンビナントの作成は,酵素の大量発現・精製とin vitro における解析などに有用である。
生化学研究室(旧生体機能化学研究室を含む)ではこれまでに数多くのセラミド/スフィンゴ脂質関連遺伝子を同定してきた。これらは,セラミド/スフィンゴ脂質の合成あるいは修飾,長鎖塩基/長鎖塩基1-リン酸の分解(詳細は「2. 長鎖塩基の代謝と脂肪酸α酸化経路の解明」を参照),極長鎖脂肪酸伸長(詳細は「3. 極長鎖脂肪酸の産生,生理機能,病態」を参照),アシルセラミド合成(詳細は「4. セラミドによる皮膚バリア形成」を参照)において働く哺乳類 16 遺伝子(ELOVL1,HACD1/2,KDSR,CERS3/6,FADS3,CYP4F22,FATP4,PNPLA1,ABHD5,SGPP2,ALDH3A2/B2,TECR,HACL2)と酵母 6 遺伝子(CSH1,RSB1,HFD1,FAA1/4,MPO1)である。図4にセラミド代謝経路と生化学研究室で同定した遺伝子群(赤,哺乳類遺伝子;青,酵母遺伝子)を示す。
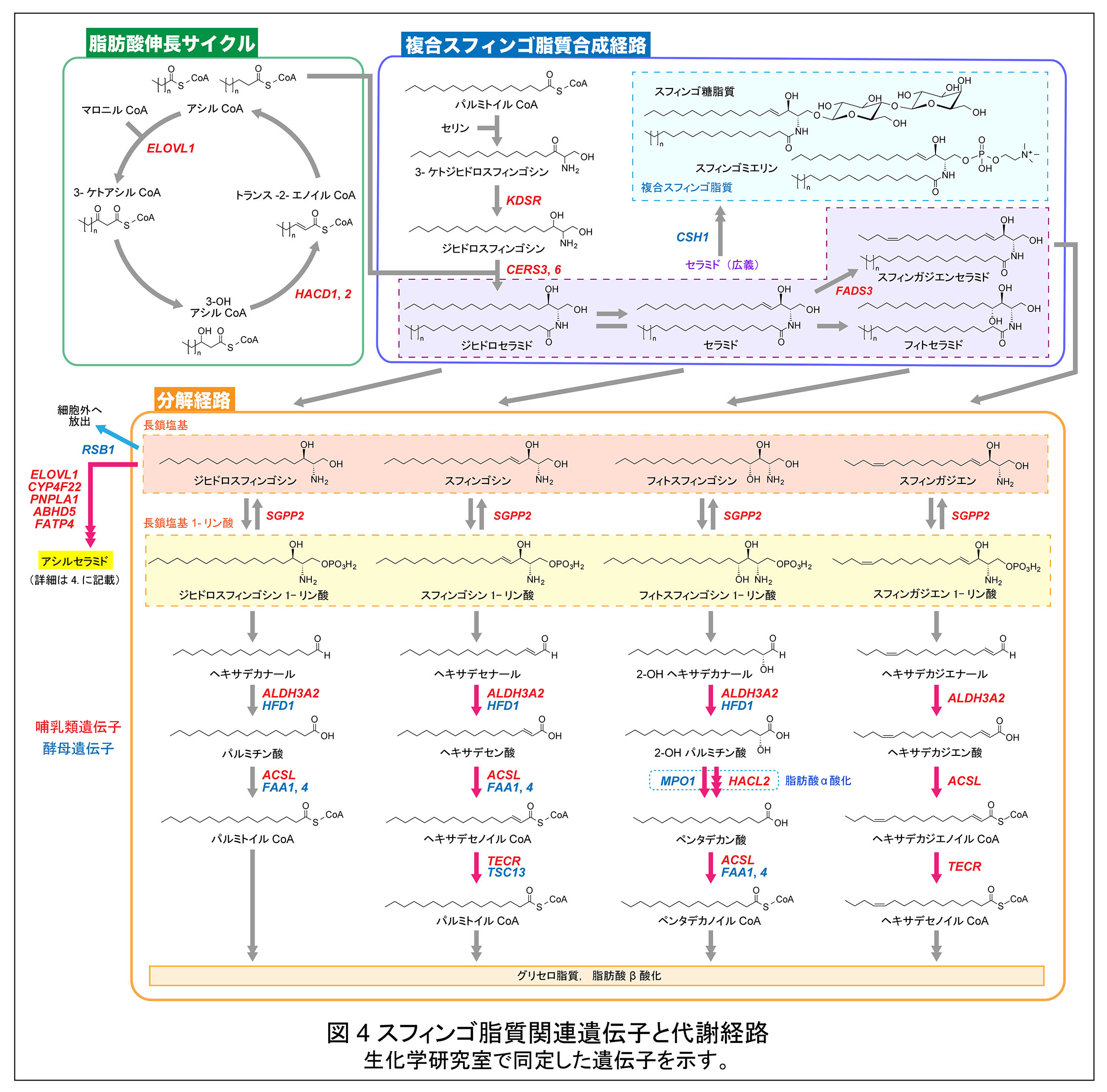
生化学研究室で同定・機能解明した遺伝子
セラミド合成/修飾関連
- 3-ケトジヒドロスフィンゴシン還元酵素 KDSR (哺乳類): スフィンゴ脂質合成の第二段階である 3-ケトジヒドロスフィンゴシンからジヒドロスフィンゴシンへの変換を担う還元酵素の遺伝子として KDSR (別名 FVT-1)を同定した1)。KDSR 遺伝子変異が道化師様魚鱗癬を引き起こすことを見出した2)。
- セラミド不飽和化酵素 FADS3 (哺乳類): スフィンガジエン(4,14-スフィンガジエン)に特徴的な 14 位シス二重結合を導入する不飽和化酵素の遺伝子としてFADS3 を同定した3)。その後の研究で,FADS3 の基質が長鎖塩基ではなくセラミドであることなど,酵素学的性質および代謝経路の詳細を解明した4)。
- セラミド合成酵素 CERS3 (哺乳類): CERS3 (別名 LASS3)がセラミド合成酵素をコードすることを初めて示すとともに5),ケラチノサイトの分化依存的に発現が上昇することを見出して皮膚におけるセラミド産生に重要なことを示唆した6)。CERS3 変異をもつ魚鱗癬患者の解析によって,アシルセラミド量の低下が魚鱗癬の発症を引き起こすことを明らかにした7)。
- セラミド合成酵素 CERS6 (哺乳類): CERS6 (別名 LASS6)がセラミド合成酵素をコードすることを初めて示すとともに,CERS6 が C16:0 アシル CoA に高い基質特異性を示すことを明らかにした8)。
長鎖塩基/長鎖塩基1-リン酸分解経路関連
- アルデヒドデヒドロゲナーゼ ALDH3A2, ALDH3B2(哺乳類)/HFD1(酵母):長鎖塩基 1-リン酸の S1P リアーゼによる分解産物である長鎖アルデヒドを長鎖脂肪酸へ変換するアルデヒドデヒドロゲナーゼの遺伝子としてALDH3A2, B2/Hfd1 を同定した9, 10)。その同定過程では,酵母の全てのアルデヒドデヒドロゲナーゼ遺伝子欠損株の中で HFD1 の欠損株(hfd1Δ )のみ長鎖塩基分解不全を示すこと,さらにその哺乳類ホモログである ALDH3A2 が欠損した哺乳類培養細胞中でも同様に長鎖塩基分解が停止することを示した9)。ALDH3A2 と同じ ALDH3 ファミリーに属する ALDH3B2 はヒトでは偽遺伝子だが,マウスでは Aldh3a2 とともに長鎖塩基分解経路において働くことを,Aldh3a2 KO マウスや Aldh3a2 Aldh3b2 二重 KO マウスの解析から明らかにした11, 12)。ALDH3A2 は神経皮膚疾患(症候群性魚鱗癬)であるシェーグレン・ラルソン症候群(SLS)の原因遺伝子である。SLS 患者角質層のセラミドプロファイリングを行い,アシルセラミド量の低下が魚鱗癬皮膚病態を引き起こすことを解明した13)。
- アシル CoA 合成酵素 ACSL1,3,4,5,6(哺乳類)/FAA1, FAA4(酵母):ALDH3A2, B2/Hfd1 によって産生された長鎖脂肪酸をアシル CoA へと変換するアシル CoA 合成酵素の遺伝子として ACSL1–6/Faa1, 4 を同定した9, 14)。その同定過程においては,酵母の各アシル CoA 合成酵素遺伝子の単独あるいは二重欠損株を用いた解析から faa1Δ faa4Δ 株において長鎖塩基の分解経路が遮断されること9),および哺乳類培養細胞において ACSL 阻害剤処理により同様の結果が得られること14)を明らかにした。
- トランス-2-エノイル CoA 還元酵素 TECR(哺乳類):スフィンゴシン/スフィンゴシン1-リン酸の分解経路で生じたヘキサデセノイル CoA (トランス-2-ヘキサデセノイル CoA )を飽和化し,パルミトイル CoA を産生する還元酵素の遺伝子として TECR(別名 TER)を同定した15)。
- 2-ヒドロキシ脂肪酸ジオキシゲナーゼ MPO1(酵母):フィトスフィンゴシンの代謝過程において生じた 2-ヒドロキシ脂肪酸を1炭素短い脂肪酸へ変換(脂肪酸α 酸化)する過程に関わる遺伝子として MPO1 を同定した16)。その同定においては,機能未知の小胞体タンパク質をコードする遺伝子の欠損株のスクリーニングを行った。MPO1 はMetabolism of Phytosphingosine to Odd-numbered fatty acids から命名した。その後の解析から,Mpo1 が鉄イオンを捕因子とするジオキシゲナーゼであることを解明した17)。IUBMB に EC 1.14.18.12 として登録した。
- 2-ヒドロキシアシル CoA リアーゼ HACL2(哺乳類):フィトスフィンゴシンの代謝過程および脂肪酸の 2-水酸化によって生じた 2-水酸化脂肪酸のα酸化において働く 2-ヒドロキシアシル CoA リアーゼの遺伝子として HACL2 を同定した18)。その後,HACL2 と相同性を示す HACL1 が分岐鎖脂肪酸のα酸化に対して主要に働くのに対し,HACL2 は 2-水酸化脂肪酸のα酸化において主要に働くことを示した19)。
脂肪酸伸長サイクル関連
- 脂肪酸伸長酵素(エロンガーゼ)ELOVL1(哺乳類):脂肪酸伸長サイクルの第一段階のアシル CoA とマロニル CoA の縮合反応において,ELOVL1 が C20–C22 アシル CoA を C24–C26 アシル CoA へ伸長させる活性をもつことを解明した20)。さらに,ELOVL1 が皮膚において超長鎖セラミドやアシルセラミド産生を通じて皮膚バリア形成に関与すること21),涙液において極長鎖マイバム脂質の産生に関与してドライアイを防止すること22) (プレスリリース原稿),神経系において極長鎖スフィンゴ脂質(特にガラクトシルセラミドやスルファチド)の産生を通じてミエリン機能に関与することを明らかにした23)。ELOVL1 の変異が神経皮膚疾患(症候群性魚鱗癬)である IKSHD 症候群を引き起こすことを見出し24),IKSHD 症候群患者の角質層のセラミドプロファイリングによってアシルセラミド量の低下が魚鱗癬発症の主要な原因であることを明らかにした25)。
- 3-ヒドロキシアシル CoA 脱水酵素 HACD1,HACD2(哺乳類):脂肪酸伸長サイクルの第三段階の脱水反応である 3-ヒドロキシアシル CoA からトランス-2-エノイル CoA の産生を担う遺伝子として HACD1 と HACD2 を同定し26),3-Hydroxy Acyl-CoA Dehydratase から HACD と命名した。ヒトにおいて HACD2 の変異がミオパチーを引き起こすことを初めて報告した27)。
アシルセラミド合成経路関連
- 脂肪酸ω-水酸化酵素 CYF4F22(ヒト)/Cyp4f39(マウス):C30–C36 の鎖長をもつ超長鎖脂肪酸のω末端を水酸化する脂肪酸ω-水酸化酵素をコードし,アシルセラミド(皮膚;プレスリリース原稿)や OAHFA(涙液油層;プレスリリース原稿)などのω-水酸化脂質の産生に関わる遺伝子としてシトクローム P450 の一種 CYP4F22/39 を同定した28, 29)。CYP4F22 は常染色体潜性先天性魚鱗癬の原因遺伝子であり,魚鱗癬患者で見られる各種ミスセンス変異によってω水酸化活性が低下すること30),患者の角質層ではアシルセラミド量が大幅に減少していることを示した31)。
- トランスアシラーゼ PNPLA1(哺乳類):アシルセラミド産生経路において,トリグリセリド中のリノール酸をω-水酸化セラミドへ転移するトランスアシラーゼ遺伝子として PNPLA1 を同定し32)(プレスリリース原稿),IUBMB に EC 1.14.18.12 として登録した。PNPLA1 は常染色体潜性先天性魚鱗癬の原因遺伝子であり,魚鱗癬患者で見られる各種ミスセンス変異によってトランスアシラーゼ活性が低下すること33),患者の角質層ではアシルセラミドと結合型セラミドの量が大幅に減少していることを示した34)。
- PNPLA1 の反応促進因子 ABHD5(哺乳類):トランスアシラーゼである PNPLA1 の反応を促進するタンパク質をコードする遺伝子として ABHD5 を同定した35)。ABHD5 は PNPLA1 を脂肪滴へリクルートすることで,基質であるトリグリセリドの利用を促進し,その結果,PNPLA1 によるアシルセラミド産生を亢進させる35)。症候群性魚鱗癬であるシャナリン・ドルフマン症候群の原因遺伝子として知られる ABHD5 において,疾患関連ミスセンス変異を有する変異体は,PNPLA1 に対するアシルセラミド産生促進作用を示さないことを明らかにした35)。
- アシル CoA 合成酵素 FATP4(哺乳類):アシルセラミド産生経路において,CYP4F22/39 が産生したω-水酸化超長鎖脂肪酸をアシル CoA へ変換するアシル CoA 合成酵素の遺伝子としてFATP4(別名 ACSVL4, SLC27A4)を同定した36)(プレスリリース原稿)。
その他
- 長鎖塩基トランスポーター RSB1(酵母):酵母を用いた遺伝学(培地中に添加した長鎖塩基に対して耐性を示すマルチコピーサプレッサーの単離)によって長鎖塩基を細胞外へ放出するトランスポーター/トランスロカーゼ遺伝子 RSB1 を同定した37)。RSB1 は Resistance to Sphingoid Base から命名した。その後の解析により,形質膜の脂質非対称性に変化が生じると RSB1 の発現が誘導されること,そのシグナル伝達には Rim101 経路が関与する(Rim21 が脂質非対称センサー)ことを解明した38–40)。
- スフィンゴシン 1-リン酸ホスファターゼ SGPP2(哺乳類):スフィンゴシン 1-リン酸を脱リン酸化し,スフィンゴシンへ変換するホスファターゼの遺伝子として SGPP2(別名 SPP2)を同定した41)。
- IPC マンノシルトランスフェラーゼ CSH1(酵母):イノシトールホスホリルセラミド(IPC)にマンノースを転移し,マンノースイノシトールホルホリルセラミド(MIPC)を産生する糖転移酵素の遺伝子として CSH1 を同定した42)。CSH1 は CSG1 Homology から命名した。
研究業績
- Kihara A, Igarashi Y. (2004) FVT-1 is a mammalian 3-ketodihydrosphingosine reductase with an active site that faces the cytosolic side of the endoplasmic reticulum membrane. J Biol Chem, 279, 49243–49250.
- Takeichi T, Torrelo A, Lee J Y W, Ohno Y, Lozano M L, Kihara A, Liu L, Yasuda Y, Ishikawa J, Murase T, Rodrigo A B, Fernández-Crehuet P, Toi Y, Mellerio J, Rivera J, Vicente V, Kelsell D P, Nishimura Y, Okuno Y, Kojima D, Ogawa Y, Sugiura K, Simpson M A, McLean W H I, Akiyama M, McGrath J A. (2017) Biallelic mutations in KDSR disrupt ceramide synthesis and result in a spectrum of keratinization disorders associated with thrombocytopenia. J Invest Dermatol, 137, 2344–2353.
- Jojima K, Edagawa M, Sawai M, Ohno Y, Kihara A. (2020) Biosynthesis of the anti-lipid-microdomain sphingoid base 4,14-sphingadiene by the ceramide desaturase FADS3. FASEB J, 34, 3318–3335.
- Jojima K, Kihara A. (2023) Metabolism of sphingadiene and characterization of the sphingadiene-producing enzyme FADS3. Biochim Biophys Acta Mol Cell Biol Lipids, 1868, 159335.
- Mizutani Y, Kihara A, Igarashi Y. (2006) LASS3 (longevity assurance homologue 3) is a mainly testis-specific (dihydro)ceramide synthase with relatively broad substrate specificity. Biochem J, 398, 531–538.
- Mizutani Y, Kihara A, Chiba H, Tojo H, Igarashi Y. (2008) 2-Hydroxy-ceramide synthesis by ceramide synthase family: enzymatic basis for the preference of FA chain length. J Lipid Res, 49, 2356–2364.
- Yamamoto M, Sassa T, Kyono Y, Uemura H, Kugo M, Hayashi H, Imai Y, Yamanishi K, Kihara A. (2021) Comprehensive stratum corneum ceramide profiling reveals reduced acylceramides in ichthyosis patient with CERS3 mutations. J Dermatol, 48, 447–456.
- Mizutani Y, Kihara A, Igarashi Y. (2005) Mammalian Lass6 and its related family members regulate synthesis of specific ceramides. Biochem J, 390, 263–271.
- Nakahara K, Ohkuni A, Kitamura T, Abe K, Naganuma T, Ohno Y, Zoeller R A, Kihara A. (2012) The Sjögren-Larsson syndrome gene encodes a hexadecenal dehydrogenase of the sphingosine 1-phosphate degradation pathway. Mol Cell, 46, 461–471.
- Kitamura T, Takagi S, Naganuma T, Kihara A. (2015) Mouse aldehyde dehydrogenase ALDH3B2 is localized to lipid droplets via two C-terminal tryptophan residues and lipid modification. Biochem J, 465, 79–87.
- Naganuma T, Takagi S, Kanetake T, Kitamura T, Hattori S, Miyakawa T, Sassa T, Kihara A. (2016) Disruption of the Sjögren-Larsson syndrome gene Aldh3a2 in mice increases keratinocyte growth and retards skin barrier recovery. J Biol Chem, 291, 11676–11688.
- Nojiri K, Fudetani S, Arai A, Kitamura T, Sassa T, Kihara A. (2021) Impaired skin barrier function due to reduced ω-O-acylceramide levels in a mouse model of Sjögren-Larsson syndrome. Mol Cell Biol, 41, e0035221.
- Arai A, Takeichi T, Wakamoto H, Sassa T, Ito Y, Murase Y, Ogi T, Akiyama M, Kihara A. (2022) Ceramide profiling of stratum corneum in Sjögren-Larsson syndrome. J Dermatol Sci, 107, 114–122.
- Ohkuni A, Ohno Y, Kihara A. (2013) Identification of acyl-CoA synthetases involved in the mammalian sphingosine 1-phosphate metabolic pathway. Biochem Biophys Res Commun, 442, 195–201.
- Wakashima T, Abe K, Kihara A. (2014) Dual functions of the trans-2-enoyl-CoA reductase TER in the sphingosine 1-phosphate metabolic pathway and in fatty acid elongation. J Biol Chem, 289, 24736–24748.
- Kondo N, Ohno Y, Yamagata M, Obara T, Seki N, Kitamura T, Naganuma T, Kihara A. (2014) Identification of the phytosphingosine metabolic pathway leading to odd-numbered fatty acids. Nat Commun, 5, 5338.
- Seki N, Mori K, Kitamura T, Miyamoto M, Kihara A. (2019) Yeast Mpo1 is a novel dioxygenase that catalyzes the α-oxidation of a 2-hydroxy fatty acid in an Fe2+-dependent manner. Mol Cell Biol, 39, e00428-00418.
- Kitamura T, Seki N, Kihara A. (2017) Phytosphingosine degradation pathway includes fatty acid α-oxidation reactions in the endoplasmic reticulum. Proc Natl Acad Sci U S A, 114, E2616–E2623.
- Mori K, Naganuma T, Kihara A. (2023) Role of 2-hydroxy acyl-CoA lyase HACL2 in odd-chain fatty acid production via α-oxidation in vivo. Mol Biol Cell, 34, ar85.
- Ohno Y, Suto S, Yamanaka M, Mizutani Y, Mitsutake S, Igarashi Y, Sassa T, Kihara A. (2010) ELOVL1 production of C24 acyl-CoAs is linked to C24 sphingolipid synthesis. Proc Natl Acad Sci U S A, 107, 18439–18444.
- Sassa T, Ohno Y, Suzuki S, Nomura T, Nishioka C, Kashiwagi T, Hirayama T, Akiyama M, Taguchi R, Shimizu H, Itohara S, Kihara A. (2013) Impaired epidermal permeability barrier in mice lacking Elovl1, the gene responsible for very-long-chain fatty acid production. Mol Cell Biol, 33, 2787–2796.
- Sassa T, Tadaki M, Kiyonari H, Kihara A. (2018) Very long-chain tear film lipids produced by fatty acid elongase ELOVL1 prevent dry eye disease in mice. FASEB J, 32, 2966–2978.(プレスリリース原稿)
- Isokawa M, Sassa T, Hattori S, Miyakawa T, Kihara A. (2019) Reduced chain length in myelin sphingolipids and poorer motor coordination in mice deficient in the fatty acid elongase Elovl1. FASEB BioAdvances, 1, 747–759.
- Mueller N, Sassa T, Morales-Gonzalez S, Schneider J, Salchow D J, Seelow D, Knierim E, Stenzel W, Kihara A, Schuelke M. (2019) De novo mutation in ELOVL1 causes ichthyosis, acanthosis nigricans, hypomyelination, spastic paraplegia, high frequency deafness and optic atrophy. J Med Genet, 56, 164–175.
- Takahashi T, Mercan S, Sassa T, Akcapınar G B, Yararbas K, Susgun S, Iseri S A U, Kihara A, Akcakaya N H. (2022) Hypomyelinating spastic dyskinesia and ichthyosis caused by a homozygous splice site mutation leading to exon skipping in ELOVL1. Brain Dev, 44, 391–400.
- Ikeda M, Kanao Y, Yamanaka M, Sakuraba H, Mizutani Y, Igarashi Y, Kihara A. (2008) Characterization of four mammalian 3-hydroxyacyl-CoA dehydratases involved in very long-chain fatty acid synthesis. FEBS Lett, 582, 2435–2440.
- Muhammad E, Reish O, Ohno Y, Scheetz T, Deluca A, Searby C, Regev M, Benyamini L, Fellig Y, Kihara A, Sheffield V C, Parvari R. (2013) Congenital myopathy is caused by mutation of HACD1. Hum Mol Genet, 22, 5229–5236.
- Ohno Y, Nakamichi S, Ohkuni A, Kamiyama N, Naoe A, Tsujimura H, Yokose U, Sugiura K, Ishikawa J, Akiyama M, Kihara A. (2015) Essential role of the cytochrome P450 CYP4F22 in the production of acylceramide, the key lipid for skin permeability barrier formation. Proc Natl Acad Sci U S A, 112, 7707–7712.(プレスリリース原稿)
- Miyamoto M, Sassa T, Sawai M, Kihara A. (2020) Lipid polarity gradient formed by ω-hydroxy lipids in tear film prevents dry eye disease. eLife, 9, e53582.(プレスリリース原稿)
- Nohara T, Ohno Y, Kihara A. (2021) Impaired production of the skin barrier lipid acylceramide by CYP4F22 ichthyosis mutations. J Dermatol Sci, 101, 69–71.
- Takeichi T, Ohno Y, Tanahashi K, Ito Y, Shiraishi K, Utsunomiya R, Yoshida S, Ikeda K, Nomura H, Morizane S, Sayama K, Ogi T, Muro Y, Kihara A, Akiyama M. (2022) Ceramide analysis in combination with genetic testing may provide a precise diagnosis for self-healing collodion babies. J Lipid Res, 63, 100308.
- Ohno Y, Kamiyama N, Nakamichi S, Kihara A. (2017) PNPLA1 is a transacylase essential for the generation of the skin barrier lipid ω-O-acylceramide. Nat Commun, 8, 14610.(プレスリリース原稿)
- Nohara T, Ohno Y, Kihara A. (2022) Impaired production of skin barrier lipid acylceramides and abnormal localization of PNPLA1 due to ichthyosis-causing mutations in PNPLA1. J Dermatol Sci, 107, 89–94.
- Kato S, Takeichi T, Jojima K, Kato M, Omi M, Tanahashi K, Otsuka A, Muro Y, Kihara A, Akiyama M. (2025) The first reported Japanese case of PNPLA1-nonsyndromic epidermal differentiation disorder (PNPLA1-nEDD) associated with an unreported 92-base-pair duplication variant. Exp Dermatol, 34, e70130.
- Ohno Y, Nara A, Nakamichi S, Kihara A. (2018) Molecular mechanism of the ichthyosis pathology of Chanarin-Dorfman syndrome: Stimulation of PNPLA1-catalyzed ω-O-acylceramide production by ABHD5. J Dermatol Sci, 92, 245–253.
- Yamamoto H, Hattori M, Chamulitrat W, Ohno Y, Kihara A. (2020) Skin permeability barrier formation by the ichthyosis-causative gene FATP4 through formation of the barrier lipid ω-O-acylceramide. Proc Natl Acad Sci U S A, 117, 2914–2922.(プレスリリース原稿)
- Kihara A, Igarashi Y. (2002) Identification and characterization of a Saccharomyces cerevisiae gene, RSB1, involved in sphingoid long-chain base release. J Biol Chem, 277, 30048–30054.
- Kihara A, Igarashi Y. (2004) Cross talk between sphingolipids and glycerophospholipids in the establishment of plasma membrane asymmetry. Mol Biol Cell, 15, 4949-4959.
- Ikeda M, Kihara A, Denpoh A, Igarashi Y. (2008) The Rim101 pathway is involved in Rsb1 expression induced by altered lipid asymmetry. Mol Biol Cell, 19, 1922–1931.
- Obara K, Yamamoto H, Kihara A. (2012) Membrane protein Rim21 plays a central role in sensing ambient pH in Saccharomyces cerevisiae. J Biol Chem, 287, 38473–38481.
- Ogawa C, Kihara A, Gokoh M, Igarashi Y. (2003) Identification and characterization of a novel human sphingosine-1-phosphate phosphohydrolase, hSPP2. J Biol Chem, 278, 1268–1272.
- Uemura S, Kihara A, Inokuchi J, Igarashi Y. (2003) Csg1p and newly identified Csh1p function in mannosylinositol phosphorylceramide synthesis by interacting with Csg2p. J Biol Chem, 278, 45049–45055.