Research
研究内容
6. 多様な皮脂脂質によるバリア形成と毛の保持
皮脂は皮膚の保湿,撥水,抗菌,体温維持,毛の保持など多様な役割を担っており,その分泌量や組成の変化はニキビ(ざ瘡),乾燥肌,脂漏性皮膚炎,脱毛症などの皮膚疾患の発症と密接に関わっている。マイバム脂質を産生するマイボーム腺は脂腺の一種であり,その構造や機能は皮脂腺と類似している。そこで生化学研究室では,マイバム脂質研究において確立した LC-MS/MS による解析手法を利用し,マウス皮脂脂質の詳細な解析を行った。
「5. マイバム脂質によるドライアイ防止」で述べたように,マウスのマイバム脂質には,コレステリルエステル,ワックスモノエステル,ワックスジエステル(タイプ 1ω,タイプ 2α,タイプ 2ω),OAHFA,コレステリル OAHFA が存在する。一方,マウスの皮脂には,これらのうちタイプ 1ω ワックスジエステルを除く脂質クラスが存在していることが明らかになった(OAHFA のみ未測定)1)(プレスリリース原稿)。このように,マイバム脂質と皮脂脂質に含まれる脂質クラスの種類自体はおおむね共通していたが,それらを構成する分子種,すなわち脂肪酸および脂肪族アルコールの組み合わせに関しては,特に炭素鎖長の点で顕著な差異が認められた。
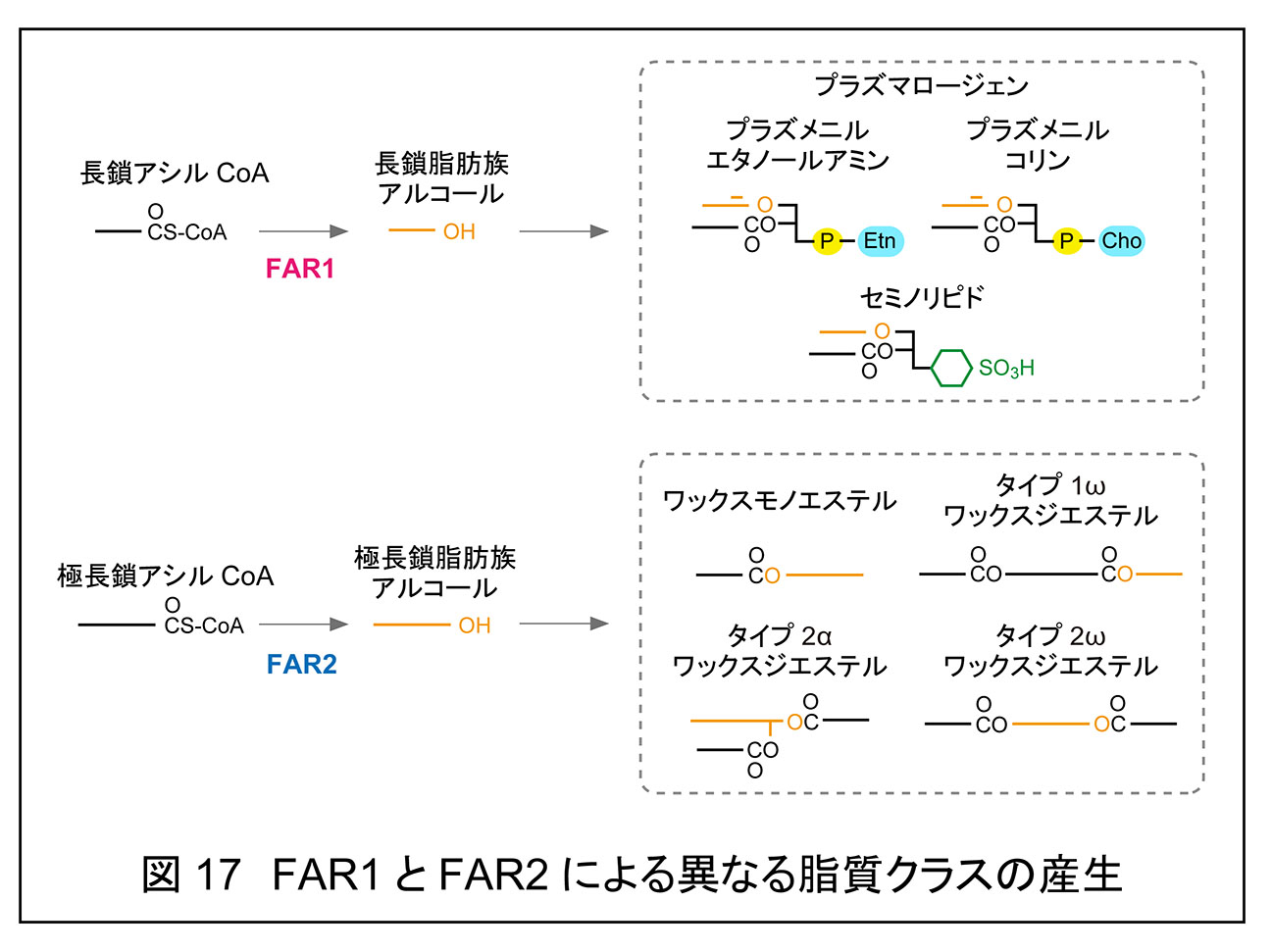
全体として,皮脂脂質はマイバム脂質と比べて分子種の多様性に富んでおり,一価不飽和の炭化水素鎖を多く含んでいた(図17)。例えば,ワックスモノエステル(脂肪酸と脂肪族アルコールのエステル体)において,マイバム脂質では脂肪酸部分として C16:1,C18:1,C16:0,脂肪族アルコール部分として C26:0,C27:0 が主要成分であったのに対し,皮脂脂質では脂肪酸部分が C15:0–C18:0 および C14:1–C22:1,脂肪族アルコール部分が C16:0–C28:0 および C31:1–C34:1 と,より広範な炭素鎖長を有し,脂肪族アルコールに一価不飽和のものが多く検出された。ワックスジエステルに関しても,特に脂肪酸部分において,皮脂脂質はマイバム脂質と比較して炭素鎖長の分布がより広範であった。
このように,皮脂とマイバムに含まれるワックスエステル(ワックスモノエステルおよびワックスジエステル)を比較すると,皮脂の方が顕著に多様性に富んでいることが分かる。これらワックスエステルの組成は,それぞれが機能する環境,特に温度条件に適応していると考えられる。マイバム脂質は涙液最外層に位置するため,角膜温度である約 32 ℃に維持されており,この温度域では液晶相をとる。一方,毛に付着する皮脂は外気温の影響を直接受けるため,幅広い温度条件に曝される。一般に,混合物の融点や相転移温度域は,その組成の多様性が高いほど広がることが知られている。皮脂中ワックスエステルの高い多様性は,このような温度変化への適応に寄与している可能性が高い。また,皮脂ワックスエステルに一価不飽和炭化水素鎖が多く含まれることは,低温環境下でも凝固を防ぐために融点を低下させる効果をもたらすと考えられる。
さらに,「5. マイバム脂質によるドライアイ防止」で述べた通り,アシル CoA 還元酵素 Far2 の KO マウスのマイバム脂質中では,ワックスモノエステルならびにタイプ 1ω,タイプ 2α,タイプ 2ω ワックスジエステルがいずれもほぼ消失していた。Far2 KO マウスはドライアイに加え,脱毛および毛の白色化(白色化は一部個体)という表現型を示しており,皮脂組成の変化が示唆されていた。実際に解析したところ,極長鎖脂肪族アルコールを含むワックスモノエステルとタイプ 2α およびタイプ 2ω ワックスジエステルが大きく減少していた。
また,アシル CoA ワックスアルコールトランスフェラーゼ Awat2 の KO マウスにおいても,一部の個体で脱毛が観察された。Awat2 KO マウスの皮脂では,長鎖脂肪酸を含むワックスモノエステルの量が野生型マウスと比較して減少しており,タイプ 2ω ワックスジエステルについても長鎖脂肪酸を含む分子種が減少していた。これらの結果は,AWAT2 が長鎖脂肪酸を含むワックスモノエステルおよびタイプ 2ω ワックスジエステルの産生を触媒することを示している。
以上のように,生化学研究室では皮脂脂質の組成を詳細に明らかにしただけでなく,それらの脂質産生に必須な遺伝子(AWAT1/2,FAR2)を同定し,その欠損が引き起こす表現型(脱毛,毛の白色化)を解明した。本研究で得られた結果および確立した解析手法は,今後,皮脂に関連する種々の病態の理解および診断への応用につながると期待される。
研究業績
- Kuribayashi K, Jojima K, Yamamoto M, Takeda M, Kihara A. (2026) Detailed composition of wax esters in mouse sebum and the involvement of FAR2 and AWAT2. iScience, 29, 114836.
(プレスリリース原稿)