Research
研究内容
3. 極長鎖脂肪酸の産生,生理機能,病態
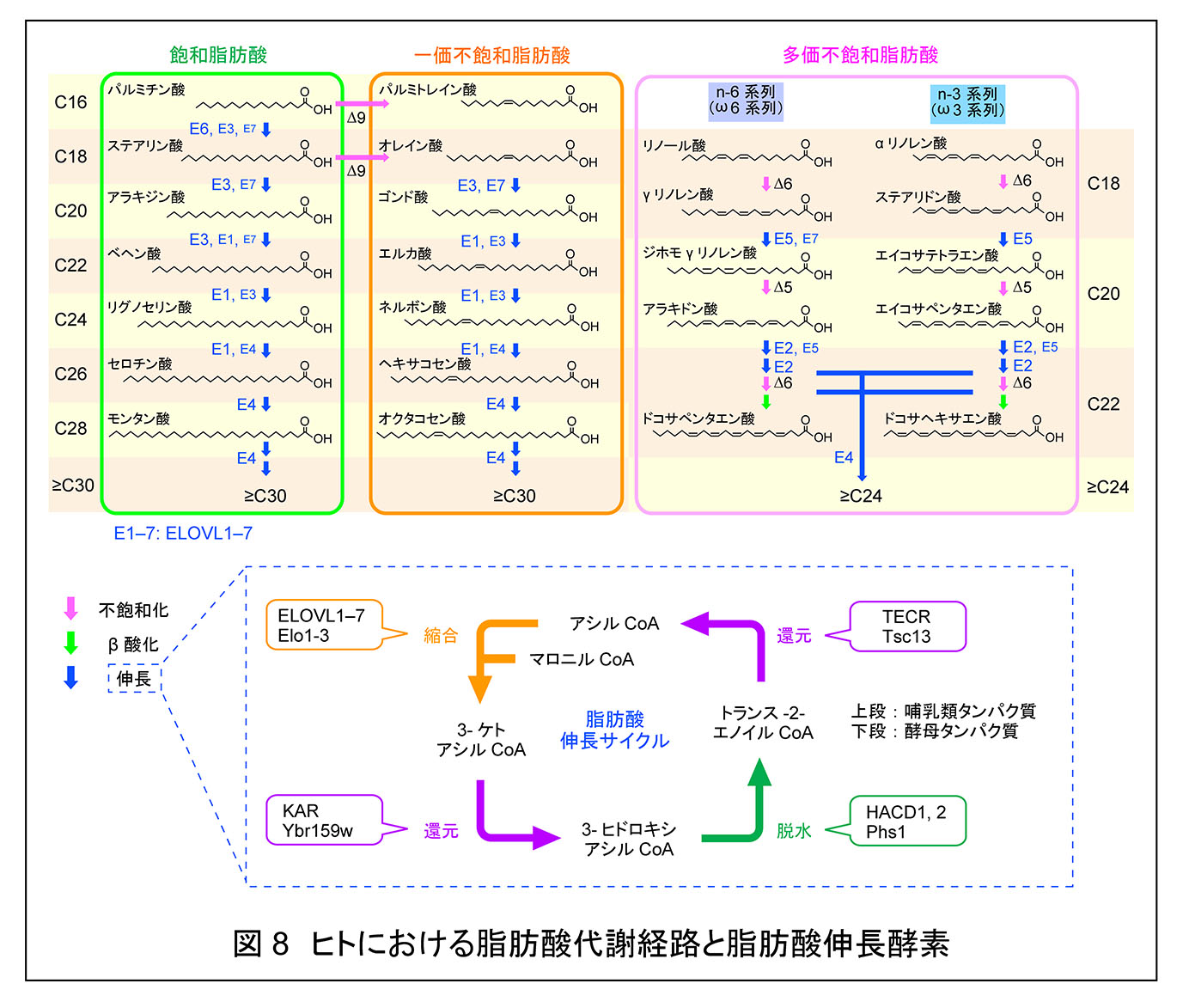
脂肪酸は炭素鎖長(C)および二重結合の数に基づいて分類される。炭素鎖長別では,脂肪酸は短鎖脂肪酸(C2–C4),中鎖脂肪酸(C5–C10),長鎖脂肪酸(C11–C20),および極長鎖脂肪酸(C21以上)に分類される。極長鎖脂肪酸のうち,C26 以上のものは,その脂肪酸が含まれる脂質クラスや性質が C21–C25 の極長鎖脂肪酸と異なることが多いため,「超長鎖脂肪酸」として記述することが有用であり,本サイトでもこの名称を用いる。一方,二重結合の数に基づく分類では,二重結合をもたないものを飽和脂肪酸,1 つ有するものを一価不飽和脂肪酸,2 つ以上有するものを多価不飽和脂肪酸と呼ぶ。なお,生体内に存在する不飽和脂肪酸の多くは,シス型をとる。脂肪酸はさまざまな脂質,すなわち,グリセロ脂質(グリセロリン脂質,トリアシルグリセロールなど),スフィンゴ脂質,ワックスエステル,コレステリルエステルなどの構成要素として利用される。それぞれの脂質を構成する脂肪酸の炭素鎖長や不飽和度は,脂質の物理化学的特性を決定する重要な要因であり,その特性は各脂質が存在する細胞や組織において求められる膜機能や膜構造に適したものとなっている。
生体膜を構成するグリセロリン脂質とスフィンゴ脂質を比較すると,グリセロリン脂質は主に C16–C20 の長鎖脂肪酸から構成されるのに対し,スフィンゴ脂質は主に C16–C24 の長鎖および極長鎖脂肪酸から構成される。このように,極長鎖脂肪酸を含有するものが多いことが,スフィンゴ脂質の特徴の一つである。また,脂肪酸の不飽和度の観点からも両者は異なる。グリセロリン脂質には飽和脂肪酸から多価不飽和脂肪酸までの幅広い脂肪酸が存在し,特に sn-2 位には不飽和脂肪酸が多く導入されている。一方,スフィンゴ脂質中の脂肪酸は主に飽和であり,不飽和脂肪酸は例外的に C24 の鎖長にのみ見られる。これらの違いはそれぞれが形成する膜の性質にも反映される。グリセロリン脂質は生体膜の基本構成要素として,適度な膜流動性を確保するために不飽和脂肪酸を多く含むのに対し,スフィンゴ脂質はコレステロールとともに流動性の低い脂質マイクロドメインを形成するため,脂質同士が密にパッキングしやすい飽和脂肪酸を主に含んでいる。
哺乳類に存在する脂肪酸のほとんどは長鎖脂肪酸であり,極長鎖脂肪酸はおそらく全体の 5% にも満たない。極長鎖脂肪酸のうちで最も広範な組織に存在するものは,スフィンゴ脂質を構成する C22 および C24 の脂肪酸である。超長鎖脂肪酸の存在割合はさらに低く,限られた器官・組織に存在する特定の脂質にのみ含まれ,それらの器官・組織の特殊な膜構造あるいは機能を支えている。これらの脂質としては,角質層脂質ラメラを形成するセラミド,涙液油層に存在するマイバム脂質,皮脂を構成する皮脂脂質,ならびに視細胞外節のディスク膜および脳の神経細胞に存在する一部のホスファチジルコリンがある1–4)。
生化学研究室では,スフィンゴ脂質の特徴である C22 および C24 の極長鎖脂肪酸に着目し,その産生に関わる脂肪酸伸長酵素(脂肪酸エロンガーゼ)ELOVL1 を同定した5)。さらに,Elovl1 ノックアウト(KO)マウスの作成と解析を行った結果,皮膚バリア異常やドライアイの表現型が見出され,これらの表現型を契機として皮膚バリアや涙液油層機能に関する研究へと展開した6, 7)。これらの詳細については,下記および「4. セラミドによる皮膚バリア形成」,「5. マイバム脂質によるドライアイ防止」を参照されたい。
各組織・器官ごとのセラミド脂肪酸組成
図8 にマウスの各組織・器官におけるセラミドの脂肪酸鎖長の解析結果を示す。本解析では,長鎖塩基にスフィンゴシン,脂肪酸に非水酸化脂肪酸をもつ最も一般的なセラミドクラス(NS)を対象とした。組織・器官ごとにセラミド中の脂肪酸組成は異なるものの,全体としては C16:0,C18:0,C22:0,C24:0,C24:1 が多くの組織・器官で主要成分を占め,これに次いで C20:0,C23:0,C24:2,C25:0 が比較的多くみられる。量的には,セラミドは多くの組織・器官でスフィンゴミエリンのおおよそ 10 分の 1 程度にとどまる。これは,セラミドが複合スフィンゴ脂質の代謝中間体として一過的に存在するためである。一方,表皮は他の組織・器官と比較して著しく多量のセラミドを含むため,本図では独立したグラフとして示した。この量の多さは,角質層においてセラミド自体が脂質ラメラの主要構成成分として機能することを反映している。また,表皮では極長鎖脂肪酸含有セラミドの割合が極めて高く(約 99 %),その中でもとりわけ C26:0 のような超長鎖脂肪酸が多く存在する。C26:0 は他の組織・器官ではほとんど認められない(食道を除く)。さらに表皮は,ここでは示していないアシルセラミドや結合型セラミドなどの特徴的なセラミドも有する(詳細は「4. セラミドによる皮膚バリア形成」を参照)。表皮以外の組織・器官における極長鎖脂肪酸の割合は 26–92 %の範囲にあり,筋肉や脳で低く,脾臓や肝臓で高い。脳には C18:0 および C24:1 を含むセラミドが多いが,これは神経細胞に C18:0 を含むスフィンゴ脂質,オリゴデンドロサイトに C24:1 を含むスフィンゴ脂質(ガラクトシルセラミド,スルファチド,スフィンゴミエリン[SM]など)が多く存在することを反映している8)。セラミド中の脂肪酸は主に飽和であるが,例外的に C24:1 は多くみられ,C24:2 も一定割合で存在し,特に脾臓で比較的高い割合を示す9)。スフィンゴ脂質は脂質マイクロドメインを形成することが知られているが,SM の脂肪酸分子種ごとの分布を解析した結果,飽和脂肪酸を含む SM と比較して,C24:1 含有 SM のドメイン内存在比は低く,C24:2 含有 SM ではさらに低いことが明らかとなった9)。これらの結果は,すべての SM が脂質マイクロドメインに局在するのではなく,特に不飽和脂肪酸含有 SM はドメイン外にも分布することを示している。
セラミドには,C23:0 や C25:0 といった奇数鎖脂肪酸をもつものが比較的多く存在する(図8)が,これは 2-水酸化脂肪酸のα酸化によって生じたものである。例えば,2-水酸化 C24:0 脂肪酸はα酸化によって(非水酸化)C23:0 脂肪酸へと変換され,C23:0 のまま,あるいは脂肪酸伸長により C25:0 へと変換された後,セラミドに取り込まれる10, 11)。生化学研究室では α 酸化の分子機構についてその大部分を解明しており,詳細は「2. 長鎖塩基の代謝と脂肪酸α酸化経路の解明」の項を参照されたい。組織特異的なセラミド中の脂肪酸としては,上記の C26:0 に加えて,精巣に特異的な C30:5 および C30:6 が存在する。これら多価不飽和の超長鎖脂肪酸を含むセラミドおよびスフィンゴミエリンは,精子形成に必須である。
脂肪酸伸長サイクル
脂肪酸の伸長は小胞体で行われる。脂肪酸はアシル CoA に変換後,縮合,還元,脱水,還元の 4 つのステップを 1 サイクルとする脂肪酸伸長サイクルによって,炭素数が 2 ずつ伸長される(図9)1, 2)。このうち最初の縮合反応が律速段階であり,脂肪酸エロンガーゼによって触媒される。哺乳類には 7 種類の脂肪酸伸長酵素(ELOVL1–7)が存在し,それぞれ異なる基質特異性を示す5)。 生化学研究室では,さまざまなアシル CoA 基質を用いたin vitro 解析により,各ELOVL の基質特異性の全体像を明らかにした(図9)5)。この解析により,ELOVL1 がスフィンゴ脂質に特徴的な C24 脂肪酸産生に対して最も高い活性を示す脂肪酸エロンガーゼであることを見出した。さらに,ELOVL1 をノックダウンした HeLa 細胞では,C24 スフィンゴ脂質が減少し,C16 スフィンゴ脂質が増加することを示した。このような細胞では,Src ファミリーキナーゼ Lyn の活性化が抑制され,アポトーシスが亢進するなど,細胞シグナル伝達に変化が生じていた5, 12)。続いて,in vivo における Elovl1 の生理的役割を解析するため,Elovl1 KO マウスを作製・解析した。その結果,この KO マウスは皮膚バリア異常により新生致死となることが明らかとなった6)。さらに,KO マウスでは表皮における C26 以上の超長鎖セラミド(アシルセラミド,結合型セラミドを含む)の減少に加え,脳や肝臓などの表皮以外の組織において C22 および C24 の極長鎖スフィンゴ脂質の減少が観察された。
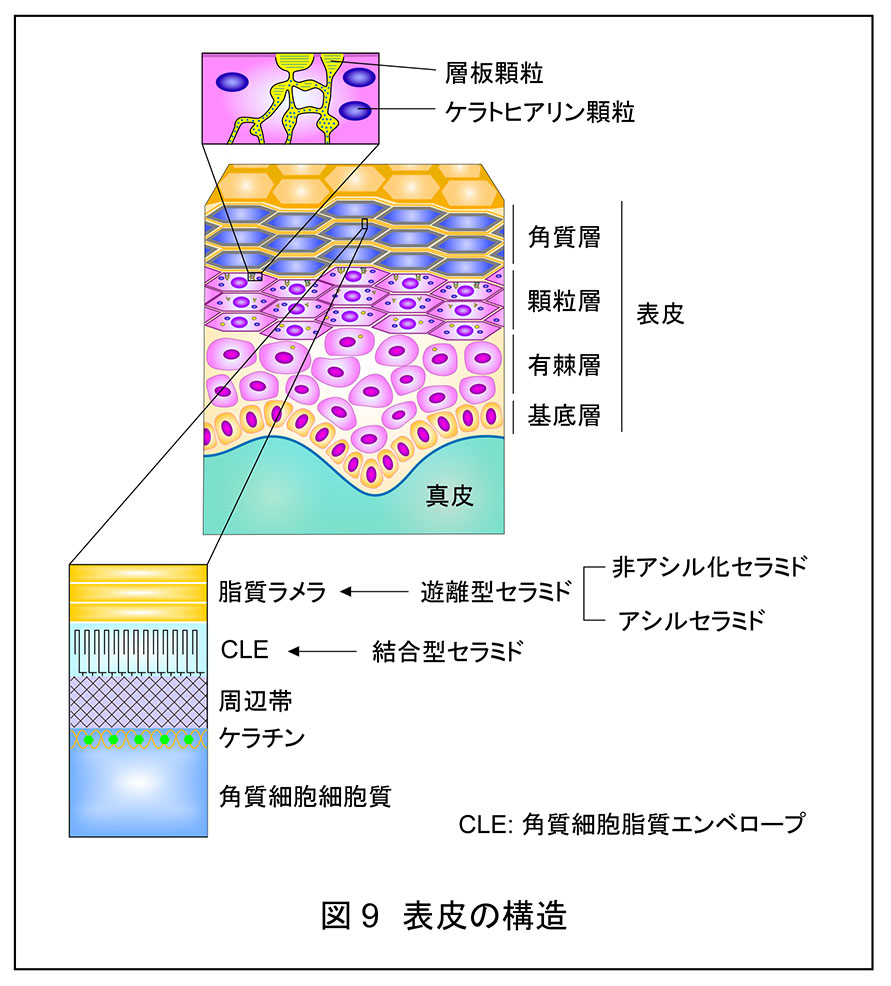
図8 の右に,各種脂肪酸に対応するセラミド合成酵素を示した。哺乳類には CERS1–6 の 6種類のセラミド合成酵素が存在し,それぞれ基質であるアシル CoA に対して異なる基質特異性を示す13–16)。すなわち,各セラミド分子種は,特定の ELOVL によるアシル CoA の伸長と,それに続く CERS による長鎖塩基とのアミド結合形成が連携して行われることによって生成される。組織・器官ごとにセラミドの脂肪酸組成が異なるのは,発現する ELOVL と CERS アイソザイムの組み合わせが異なるためである。例えば,脳に多い C18:0 セラミドは ELOVL6 と CERS1,多くの組織に見られる C24:0 および C24:1 セラミドは ELOVL1 と CERS2,表皮に存在する C26:0 セラミドは ELOVL1 と CERS3,さらに同じく表皮で見られる C28 以上のセラミドは ELOVL4 と CERS3 によって産生される17, 18)。ELOVL1 は連携する CERS の種類に応じてアシル CoA 伸長の到達鎖長が異なり,CERS2 との組み合わせでは C24 まで伸長するのに対し,CERS3 との組み合わせでは C24 で停止せず,さらに C26 まで伸長する6, 19)。また,ELOVL1 によって産生された C26 アシル CoA は ELOVL4 によってさらに伸長され,CERS3 の基質となる19)。 CERS3 の発現は表皮と精巣にほぼ限定されている14, 20)。表皮では,CERS3 と ELOVL4 は有棘層以降で発現が誘導される一方,CERS2 の発現は低下し,ELOVL1 は顆粒層で発現が上昇する6)。これらの発現変化が,表皮における特徴的なセラミド脂肪酸組成を生み出している。セラミド産生において連続的な反応を担う ELOVL と CERS の間には,CERS による ELOVL 活性の調節機構が存在することを我々は見出している。例えば,ELOVL1 および ELOVL4 の活性は,それぞれ CERS2 および CERS3 の存在下で上昇する5, 21)。ただし,その詳細な分子機構は未だ明らかではない。 CERS の活性調節機構については,CERS1 を除く CERS がリン酸化による調節を受けることを見出している15)。CERS2—6 の非リン酸化型変異体は,野生型と比べて活性が低い。これらのリン酸化は通常生育条件下で維持されているが,どのような条件で脱リン酸化が起こるかについては未解明である。
全身で Elovl1 を欠損した KO マウスは皮膚バリア形成不全により新生致死となるため6),表皮以外の組織における極長鎖脂肪酸の役割の解明には至らなかった。そこで生化学研究室では,表皮においてのみ Elovl1 の発現を回復させた Elovl1 KO マウスを作製した。このマウスは,表皮特異的に発現するインボルクリン(Inv)プロモーターの制御下で Elovl1 を発現するトランスジェニック(Tg)マウスと Elovl1 KO マウスを交配することで作製した。以降,本マウスを Tg-Elovl1 KO マウスと表記する。Tg-Elovl1 KO マウスは皮膚バリア異常を示さず,成体にまで成長したが,ドライアイを示し,5 ヶ月齢以降には角膜の混濁が観察された7)。その詳細については「5. マイバム脂質によるドライアイ防止」を参照されたい。また,Tg-Elovl1 KO マウスは運動障害およびミエリン低形成を示した22)。この変異マウスの脳では,ミエリンの機能および形態維持に重要なガラクトシルセラミドの量が減少しており,さらにガラクトシルセラミド,スルファチド,スフィンゴミエリンといった複合スフィンゴ脂質において脂肪酸鎖長の短鎖化が認められた。これらの変化が神経症状の発現に関与していると考えられる。 全身での Elovl1 KO マウスおよび Tg-Elovl1 KO マウスの解析で観察された皮膚および神経症状は,後にヒトにおいても ELOVL1 変異によって引き起こされることが明らかとなった。我々は 2019 年にドイツの研究者との共同研究により,ELOVL1 変異が皮膚魚鱗癬,下肢痙性,歩行困難,ミエリン低形成などを伴う神経皮膚疾患(IKSHD 症候群)を引き起こすことを初めて報告した23)。
極長鎖脂肪酸のβ酸化はペルオキシソームで行われる。極長鎖アシル CoA のペルオキシソームへの輸送には ABC トランスポーター ABCD1 が関与する。その遺伝子変異は極長鎖脂肪酸の蓄積を引き起こし,副腎白質ジストロフィー(X-ALD)の原因となる。ロレンツォオイルはオレイン酸(C18:1)およびエルカ酸(C22:1)のトリアシルグリセロールであるグリセリルトリオレートとグリセリルトリエルケートの約 4:1 の混合物であり,血漿中の飽和極長鎖脂肪酸(C24:0,C26:0)を低下させる効果が報告されている。しかし,その作用機序は長らく不明であった。生化学研究室では,オレイン酸とエルカ酸の遊離脂肪酸 4:1 混合物(ロレンツォオイルの加水分解物に相当)が ELOVL1 の活性を阻害することにより,細胞における飽和極長鎖脂肪酸の産生を抑制することを明らかにした24)。一方,これらの遊離脂肪酸混合物の添加は,一価不飽和極長鎖脂肪酸(C22:1,C24:1 など)の産生を増加させた。ELOVL1 はin vitro において,飽和だけでなく一価不飽和極長鎖脂肪酸の産生に対しても活性を示すが,一価不飽和極長鎖脂肪酸の産生には ELOVL3 や ELOVL7 など他の ELOVL の活性も高いため5),細胞や組織における ELOVL1 の寄与は飽和極長鎖脂肪酸の場合と比べて相対的に小さい。また,ELOVL1 の機能あるいは発現低下は,さまざまな組織において ELOVL3 やELOVL7 の代償的発現増加を引き起こす7, 25)。これらの要因が,上記脂肪酸混合物の添加によって一価不飽和極長鎖脂肪酸の産生が増加した原因であると考えられる。ロレンツォオイルは治療効果の点で課題が残されているが,我々の結果は ELOVL1 が X-ALD 治療における有力な分子標的であることを示しており,今後より選択的かつ有効な ELOVL1 阻害剤の開発が期待される。
ELOVL4 は炭素数 28 以上の超長鎖脂肪酸の産生に関わる。これらの脂肪酸を含む脂質は,表皮(アシルセラミドなど),脳および網膜(ホスファチジルコリン),精子(セラミド,スフィンゴミエリン),マイボーム腺および皮脂腺(コレステリルエステル,ワックスエステルなど)といった限られた組織・器官に存在する1–4)。ELOVL4 の変異はこれらの組織の機能および形態異常を引き起こし,黄斑変性疾患,神経疾患,皮膚魚鱗癬の発症につながる。ELOVL4 変異は C 末端欠失を伴う短縮型変異(スタルガルト病 3 型),機能改変を伴うミスセンス変異(脊髄小脳性運動失調症[SCA]34 型),および機能喪失型変異(ISQMR)に大別される。このうち SCA34 は,小脳萎縮と運動失調を特徴とする常染色体顕性遺伝性疾患である。生化学研究室では,SCA34 の顕性遺伝形式が,変異体タンパク質による超長鎖脂肪酸の不完全伸長によって超長鎖ホスファチジルコリンがより短鎖の分子に置き換わることに起因すると考えられることを示した26)。
ELOVL は膜内在性タンパク質であるため,精製には非イオン性界面活性剤(Triton X-100 など)による可溶化が必要である。しかし,そのような可溶化条件下では ELOVL は活性を示さない。生化学研究室では,可溶化した ELOVL7 を精製後,プロテオリポソームに再構成することでこの問題を回避し,活性測定に成功した27)。ELOVL7 の基質特異性は,過剰発現膜を用いた活性測定の結果と一致して5),C18–C20 のアシル CoA に対して高かった27)。さらに,我々は C16:0-CoA に活性を示す ELOVL6 についてもプロテオリポソームに再構成し,活性を測定した。その結果,脂肪酸伸長サイクルの第二段階である還元反応を触媒する KAR を ELOVL6 とともに再構成すると,ELOVL6 の活性が単独の場合よりも上昇することを見出した28)。このことは,脂肪酸伸長サイクルの第一段階と第二段階が協調して進行することを示している。
生化学研究室では,3-ヒドロキシアシル CoA 脱水酵素として HACD1–4 を同定した29)。これらはいずれも in vitro において 3-ヒドロキシパルミトイル CoA に対して活性を示した。 一方,これらの HACD 遺伝子をノックアウトした細胞を用いた解析から,HACD1 および HACD2 が幅広い脂肪酸伸長反応に関与することが示されたのに対し,HACD3 および HACD4 の関与は細胞レベルでは小さいか,ほとんど認められなかった。 HACD2 は広範な組織・器官で発現し,それらにおける主要な酵素として機能するのに対し,HACD1 は心臓や骨格筋などの筋組織に特異的に発現して機能する。我々はイスラエルの研究者との共同研究により,HACD1 変異がヒトにおいてミオパチーを引き起こすことを初めて報告した30)。また,同様に Hacd1 KOマウスもミオパチーの表現型を示す31)。
脂肪酸伸長サイクルの第四段階を触媒する TECR の p.P182L ミスセンス変異は非症候群性の知的障害を引き起こすことが知られている。我々はこの変異によってタンパク質の安定性および酵素活性が低下すること,ならびに患者由来の B リンパ芽球様細胞株において C24 極長鎖脂肪酸含有スフィンゴ脂質量が低下することを明らかにした32)。また,TECR の存在量の低下が脂肪酸伸長サイクル第三段階を担う HACD の活性低下を引き起こすことを見出した。このことは,脂肪酸伸長サイクルの第一段階と第二段階に加えて,第三段階と第四段階も協調して機能していることを示している。TECR P182L 変異タンパク質は活性を完全には失っておらず,脂肪酸伸長サイクルへの影響は比較的軽微である。それにもかかわらず神経系に異常が生じることは,神経系における極長鎖脂肪酸の重要性が他の組織よりも高いことを示唆している。ミエリンは神経細胞の軸索を被覆し,絶縁体として機能する。ミエリンは脂質に富み,その絶縁性は主として脂質によって付与されている。とりわけ,C24 鎖長をもつガラクトシルセラミドやスルファチドなどの極長鎖スフィンゴ脂質はミエリンの形成および機能維持に不可欠であり,TECR の p.P182L 変異はこれらの脂質量の低下を介してミエリン機能を低下させると考えられる。後述するように,極長鎖脂肪酸はミエリンのような曲率の高い膜構造を安定化していると予想される。
極長鎖脂肪酸は酵母からヒトに至るまで真核生物に広く存在している。出芽酵母 Saccharomyces cerevisiae における極長鎖脂肪酸はほぼ全てが C26 であり,スフィンゴ脂質中に存在する。極長鎖脂肪酸の合成が不全となった酵母は致死となる。脂肪酸伸長サイクルの 4 つの反応を担う酵素はいずれも真核生物で保存されており,酵母 Phs1 は哺乳類 HACD1–4 のホモログである。生化学研究室では Phs1 の活性に重要な残基を同定し,反応機構に関する知見を得た33, 34)。また,極長鎖脂肪酸の細胞レベルでの生理機能を明らかにすることを目的として,酵母を用いた遺伝学的解析を行った。極長鎖脂肪酸の産生量が低下すると酵母は温度感受性を示す。そこで,この性質を利用し,温度感受性をレスキューするマルチコピーサプレッサーの単離を行った。その結果,後期エンドソーム(マルチベシキュラーボディー;MVB)を介した小胞輸送に関与する VPS21 を同定した35)。この結果は,極長鎖脂肪酸を含むスフィンゴ脂質が MVB を介した小胞輸送において重要な機能を果たすことを示唆している。極長鎖脂肪酸はその長さゆえに脂質二重層の両層にまたがって局在することで,曲率の高い不安定な膜構造の安定化に寄与すると考えられる。MVB では内腔小胞形成の過程でこのような高曲率の膜構造が形成されるため,極長鎖脂肪酸の要求性が高いと推測される。また,上述のように TECR や ELOVL1 の変異は神経症状を引き起こす。ミエリンは軸索を取り囲む多層構造をもち,高い曲率を有する膜構造体であることから,その安定化に極長鎖スフィンゴ脂質が寄与しているのであろう。
脂肪酸伸長サイクルの第四段階を触媒するトランス-2-エノイル CoA 還元酵素(酵母 Tsc13,哺乳類 TECR)の触媒基や反応機構は不明であったが,生化学研究室では,Tsc13/TECR の Tyr256/Tyr248 が触媒残基として機能し,NADPH からのヒドリド移動により生じたエノラート中間体に対してプロトンを供与することで反応が進行するモデルを提唱した36)。さらに,酵母において Tsc13 と脂肪酸伸長サイクルの第一段階を担う脂肪酸エロンガーゼ(Elo2 および Elo3)との相互作用を明らかにし,その相互作用が脂肪酸エロンガーゼの活性発現に重要であることを示した36)。 すなわち,脂肪酸伸長サイクルにおいては,第一段階と第二段階ならびに第三段階と第四段階のみならず,第四段階と次のサイクルの第一段階も協調して進行している。
以上のように,脂肪酸伸長サイクルは各段階が相互に連携しながら進行し,極長鎖脂肪酸の産生とその鎖長制御を精密に担っている。これらの知見は,極長鎖脂肪酸が生体膜に加えて脂質ラメラやミエリンなどの膜構造の形成と機能維持を支える基盤分子であることを示すとともに,その異常が皮膚や神経系を中心とした多様な疾患の発症につながる分子機構の理解に重要な手がかりを与えるものである。
研究業績
- Kihara A. (2012) Very long-chain fatty acids: elongation, physiology and related disorders. J Biochem, 152, 387–395, Review.
- Sassa T, Kihara A. (2014) Metabolism of very long-chain fatty acids: genes and pathophysiology. Biomol Ther, 22, 83–92, Review.
- 木原章雄. (2018) 鎖長の違いによる脂肪酸クオリティ — 極長鎖・超長鎖脂肪酸の産生機構と病態. 医学のあゆみ, 264, 966–971, Review.
- 木原章雄. (2019) 極長鎖脂質による表皮および涙液における透過性バリア形成. 医学のあゆみ, 269, 983–988, Review.
- Ohno Y, Suto S, Yamanaka M, Mizutani Y, Mitsutake S, Igarashi Y, Sassa T, Kihara A. (2010) ELOVL1 production of C24 acyl-CoAs is linked to C24 sphingolipid synthesis. Proc Natl Acad Sci USA, 107, 18439–18444.
- Sassa T, Ohno Y, Suzuki S, Nomura T, Nishioka C, Kashiwagi T, Hirayama T, Akiyama M, Taguchi R, Shimizu H, Itohara S, Kihara A. (2013) Impaired epidermal permeability barrier in mice lacking Elovl1, the gene responsible for very-long-chain fatty acid production. Mol Cell Biol, 33, 2787–2796.
- Sassa T, Tadaki M, Kiyonari H, Kihara A. (2018) Very long-chain tear film lipids produced by fatty acid elongase ELOVL1 prevent dry eye disease in mice. FASEB J, 32, 2966–2978.
- Sugimoto M, Shimizu Y, Yoshioka T, Wakabayashi M, Tanaka Y, Higashino K, Numata Y, Sakai S, Kihara A, Igarashi Y, Kuge Y. (2015) Histological analyses by matrix-assisted laser desorption/ionization-imaging mass spectrometry reveal differential localization of sphingomyelin molecular species regulated by particular ceramide synthase in mouse brains. Biochim Biophys Acta, 1851, 1554–1565.
- Edagawa M, Sawai M, Ohno Y, Kihara A. (2018) Widespread tissue distribution and synthetic pathway of polyunsaturated C24:2 sphingolipids in mammals. Biochim Biophys Acta Mol Cell Biol Lipids, 1863, 1441–1448.
- Mori K, Obara T, Seki N, Miyamoto M, Naganuma T, Kitamura T, Kihara A. (2020) Catalytic residues, substrate specificity, and role in carbon starvation of the 2-hydroxy FA dioxygenase Mpo1 in yeast. J Lipid Res, 61, 1104–1114.
- Mori K, Naganuma T, Kihara A. (2023) Role of 2-hydroxy acyl-CoA lyase HACL2 in odd-chain fatty acid production via α-oxidation in vivo. Mol Biol Cell, 34, ar85.
- Sassa T, Suto S, Okayasu Y, Kihara A. (2012) A shift in sphingolipid composition from C24 to C16 increases susceptibility to apoptosis in HeLa cells. Biochim Biophys Acta, 1821, 1031–1037.
- Mizutani Y, Kihara A, Igarashi Y. (2005) Mammalian Lass6 and its related family members regulate synthesis of specific ceramides. Biochem J, 390, 263–271.
- Mizutani Y, Kihara A, Igarashi Y. (2006) LASS3 (longevity assurance homologue 3) is a mainly testis-specific (dihydro)ceramide synthase with relatively broad substrate specificity. Biochem J, 398, 531–538.
- Sassa T, Hirayama T, Kihara A. (2016) Enzyme activities of the ceramide synthases CERS2–6 are regulated by phosphorylation in the C-terminal region. J Biol Chem, 291, 7477–7487.
- 木原章雄. (2019) スフィンゴ脂質クオリティによる生体防御. The Lipid, 30, 28–34, Review.
- 木原章雄. (2015) セラミドによる皮膚バリア形成. 実験医学, 33, 117–-122, Review.
- Kihara A. (2016) Synthesis and degradation pathways, functions, and pathology of ceramides and epidermal acylceramides. Prog Lipid Res, 63, 50–69, Review.
- Ohno Y, Nakamichi S, Ohkuni A, Kamiyama N, Naoe A, Tsujimura H, Yokose U, Sugiura K, Ishikawa J, Akiyama M, Kihara A. (2015) Essential role of the cytochrome P450 CYP4F22 in the production of acylceramide, the key lipid for skin permeability barrier formation. Proc Natl Acad Sci USA, 112, 7707–7712.
- Mizutani Y, Kihara A, Chiba H, Tojo H, Igarashi Y. (2008) 2-Hydroxy-ceramide synthesis by ceramide synthase family: enzymatic basis for the preference of FA chain length. J Lipid Res, 49, 2356–2364.
- Mizutani Y, Sun H, Ohno Y, Sassa T, Wakashima T, Obara M, Yuyama K, Kihara A, Igarashi Y. (2013) Cooperative synthesis of ultra long-chain fatty acid and ceramide during keratinocyte differentiation. PLoS One, 8, e67317.
- Isokawa M, Sassa T, Hattori S, Miyakawa T, Kihara A. (2019) Reduced chain length in myelin sphingolipids and poorer motor coordination in mice deficient in the fatty acid elongase Elovl1. FASEB BioAdvances, 1, 747–759.
- Mueller N, Sassa T, Morales-Gonzalez S, Schneider J, Salchow D J, Seelow D, Knierim E, Stenzel W, Kihara A, Schuelke M. (2019) De novo mutation in ELOVL1 causes ichthyosis, acanthosis nigricans, hypomyelination, spastic paraplegia, high frequency deafness and optic atrophy. J Med Genet, 56, 164–175.
- Sassa T, Wakashima T, Ohno Y, Kihara A. (2014) Lorenzo’s oil inhibits ELOVL1 and lowers the level of sphingomyelin with a saturated very long-chain fatty acid. J Lipid Res, 55, 524–530.
- Hiranuma T, Sassa T, Kihara A. (2025) Relationship between time-dependent epidermal ceramide composition changes and skin barrier function in adult mice. Mol Biol Cell, 36, ar57.
- Tamura Y, Sassa T, Nishizawa T, Kihara A. (2023) Incomplete elongation of ultra-long-chain polyunsaturated acyl-CoAs by the fatty acid elongase ELOVL4 in spinocerebellar ataxia type 34. Mol Cell Biol, 43, 85–101.
- Naganuma T, Sato Y, Sassa T, Ohno Y, Kihara A. (2011) Biochemical characterization of the very long-chain fatty acid elongase ELOVL7. FEBS Lett, 585, 3337–3341.
- Naganuma T, Kihara A. (2014) Two modes of regulation of the fatty acid elongase ELOVL6 by the 3-ketoacyl-CoA reductase KAR in the fatty acid elongation cycle. PLoS One, 9, e101823.
- Ikeda M, Kanao Y, Yamanaka M, Sakuraba H, Mizutani Y, Igarashi Y, Kihara A. (2008) Characterization of four mammalian 3-hydroxyacyl-CoA dehydratases involved in very long-chain fatty acid synthesis. FEBS Lett, 582, 2435–2440.
- Muhammad E, Reish O, Ohno Y, Scheetz T, Deluca A, Searby C, Regev M, Benyamini L, Fellig Y, Kihara A, Sheffield V C, Parvari R. (2013) Congenital myopathy is caused by mutation of HACD1. Hum Mol Genet, 22, 5229–5236.
- Blondelle J, Ohno Y, Gache V, Guyot S, Storck S, Blanchard-Gutton N, Barthelemy I, Walmsley G, Rahier A, Gadin S, Maurer M, Guillaud L, Prola A, Ferry A, Aubin-Houzelstein G, Demarquoy J, Relaix F, Piercy R J, Blot S, Kihara A, Tiret L, Pilot-Storck F. (2015) HACD1, a regulator of membrane composition and fluidity, promotes myoblast fusion and skeletal muscle growth. J Mol Cell Biol, 7, 429–440.
- Abe K, Ohno Y, Sassa T, Taguchi R, Caliskan M, Ober C, Kihara A. (2013) Mutation for nonsyndromic mental retardation in the trans-2-enoyl-CoA reductase TER gene involved in fatty acid elongation impairs the enzyme activity and stability, leading to change in sphingolipid profile. J Biol Chem, 288, 36741–36749.
- Kihara A, Sakuraba H, Ikeda M, Denpoh A, Igarashi Y. (2008) Membrane topology and essential amino acid residues of Phs1, a 3-hydroxyacyl-CoA dehydratase involved in very long-chain fatty acid elongation. J Biol Chem, 283, 11199–11209.
- Yazawa T, Naganuma T, Yamagata M, Kihara A. (2013) Identification of residues important for the catalysis, structure maintenance, and substrate specificity of yeast 3-hydroxyacyl-CoA dehydratase Phs1. FEBS Lett, 587, 804–809.
- Obara K, Kojima R, Kihara A. (2013) Effects on vesicular transport pathways at the late endosome in cells with limited very long-chain fatty acids. J Lipid Res, 54, 831–842.
- Kato R, Takenaka Y, Ohno Y, Kihara A. (2024) Catalytic mechanism of trans-2-enoyl-CoA reductases in the fatty acid elongation cycle and its cooperative action with fatty acid elongases. J Biol Chem, 300, 105656.