Research
研究内容
7. エーテル脂質の生理機能と生合成経路
バクテリアや真核生物の脂質の多くの炭化水素鎖は脂肪酸であり,グリセロール骨格とエステル結合しているが,一部の脂質の炭化水素鎖は脂肪族アルコールであり,グリセロール骨格とエーテル結合している。このような脂質は総称してエーテル脂質と呼ばれる。
エーテル結合はエステル結合よりもはるかに加水分解に対して耐性が高く,化学的に頑強である。過酷な環境で生活することの多い古細菌の脂質がエーテル脂質であることを考えるとエーテル脂質は過酷な環境下で有利に働くことが予測される。一方で,加水分解されにくいという性質は,分解と合成のバランスによって恒常性(ホメオスタシス)を維持するという点では不利に働く可能性がある。それにも関わらず,生体にエーテル脂質が存在しているということは,エステル型脂質では代替できない役割をエーテル脂質が担っていることを強く示唆している。しかし,その生理的意義や分子メカニズムの多くは,いまだ十分に解明されていない。
哺乳類に存在するエーテル脂質としては,プラズマロージェン(プラズメニルコリンおよびプラズメニルエタノールアミン),それらのプラズマニル体,血小板活性化因子(PAF),アルキルジアシルグリセロール,アルキルトリアシルグリセロール,GPI アンカー,セミノリピドなどが知られている(図18)。この中でプラズマロージェンは量的に最も豊富であり,生体膜を構成するリン脂質の約 15–20 %を占めるとされ,特に脳(ミエリン),心臓,骨格筋に多く存在する。プラズマロージェンの産生に関与する遺伝子に変異が生じると,近位四肢短縮,先天性白内障,重度の発達遅滞を主徴とする近位型点状軟骨異形成症が発症することからも,その生理的重要性が示されている。
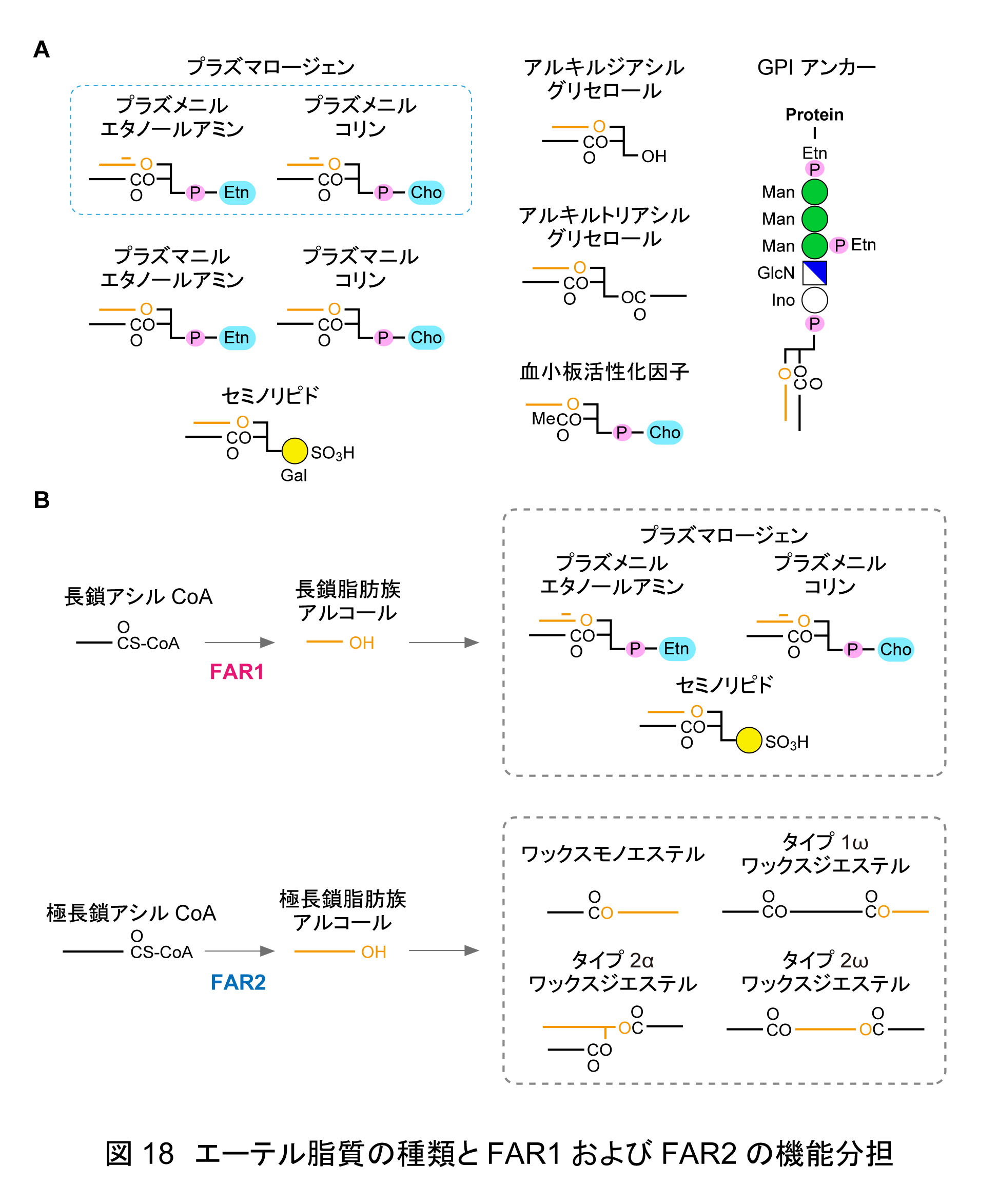
生化学研究室では,マイバム脂質の研究過程において,脂肪族アルコール産生を触媒する脂肪族アシル CoA 還元酵素 FAR1 および FAR2 に着目し,両酵素の基質特異性の違いの解明ならびに KO マウスの作製と解析を行ってきた。すでに「5. マイバム脂質によるドライアイ防止」の項で述べたように,FAR1 は主に炭素数 20 以下の長鎖脂肪族アルコールの産生に,FAR2 は炭素数 21 以上の極長鎖脂肪族アルコールの産生に関与することを我々は明らかにした1)。Far2 KO マウスはドライアイを呈し,そのマイバム中では極長鎖脂肪族アルコールを含むワックスモノエステルや,タイプ 1ω,タイプ 2α,タイプ 2ω のワックスジエステルがほぼ消失していた1)。
一方,エーテル脂質には主に長鎖脂肪族アルコールが利用されるため,その量的変動は Far2 KO マウスではなく Far1 KO マウスにおいて顕著であった。例えば,Far1 KO マウスの脳ではプラズメニルエタノールアミン量が野生型マウスの約 8 %まで低下し,これを補うようにジアシル型のホスファチジルエタノールアミンが増加していた2)。Far1 KO マウスの多くは成体まで成長できず,ミエリン形成不全や出生時の骨硬化遅延をを示した。
さらに生化学研究室では,エーテル脂質生合成経路の第 3 段階を担うアルキルグリセロンリン酸還元酵素の新規酵素として DHRS7 を同定した3)。これまで同反応を担う酵素としては DHRS7B が知られていたが,DHRS7B が主にペルオキシソームに局在するのに対し,DHRS7 は小胞体に局在していた。また,プラズメニルエタノールアミンの新規合成において,両者は異なる脂肪酸分子種の産生に関与していた。DHRS7 は脂肪酸部分が C18:1 である分子種の産生に最も高い寄与を示し,次いで C16:0 および C22:4 種の産生に寄与した一方,DHRS7B は幅広い分子種の産生に関与していた。このように,DHRS7 と DHRS7B は細胞内局在および産生するプラズマロージェン分子種の点で明確に異なる機能を担っていることが示された。
セミノリピドは精巣特異的に発現するエーテル糖脂質であり,精子形成に必須の役割を果たすことが知られている。しかし,セミノリピドに含まれるアルキル基およびアシル基の分子多様性,産生される精子形成段階,ならびにアルキル基合成に関与する FAR アイソザイム(FAR1 または FAR2 )については,これまでほとんど明らかにされていなかった。そこで,我々は Far1 KO マウスを用いてこれらの解明を行なった。Far1 は精原細胞,精母細胞,精子細胞のすべてで発現しており,雄の Far1 KO マウスは不妊であった4)。Far1 KO マウスでは精巣上体に精子が認められず,精巣は小さく,精細管内には多核細胞や空胞が観察された。LC-MS/MS 解析により,野生型マウス精巣におけるセミノリピドの 90 %以上はアルキル基,アシル基ともに C16:0 の分子種であること,さらにセミノリピドは精子形成過程のすべての細胞種に存在することが明らかとなった。これに対し,Far1 KO マウスではセミノリピドは完全に消失し,代わりにエステル型のスルホガラクトシルジアシルグリセロールが産生されていた。これらの結果から,FAR1 はセミノリピド合成および正常な精子形成に不可欠であることが示された。
以上,生化学研究室ではエーテル脂質とエステル型脂質の機能的差異を明確にするとともに,FAR1 と FAR2 の役割分担(図18),エーテル脂質生合成経路における新規酵素の同定,さらにエーテル脂質を構成する詳細な分子種とその機能を明らかにした。これらにより,エーテル脂質が分子種特異的に生体機能を制御する重要な脂質群であることが示された。
研究業績
- Otsuka K, Sawai M, Kihara A. (2022) Formation of fatty alcohols—components of meibum lipids—by the fatty acyl-CoA reductase FAR2 is essential for dry eye prevention. FASEB J, 36, e22216.(プレスリリース原稿)
- Takahashi T, Otsuka K, Sassa T, Kihara A. (2025) Acyl-CoA reductase Far1 deficiency impairs ether lipid production and hypomyelination in mouse brains. Mol Cell Biol, 45, 495-510.
- Takahashi T, Otsuka K, Sassa T, Kihara A. (2026) The acyl- and alkyl-glycerone phosphate reductase DHRS7 is involved in the production of distinct plasmalogen species from DHRS7B. J Cell Sci, 139, jcs264759.
(JCS の Research Highlight に選出) - Tamazawa A, Naganuma T, Otsuka K, Takahashi T, Sassa T, Kihara A. (2025) Fatty acyl-CoA reductase FAR1 is essential for the testicular seminolipid synthesis required for spermatogenesis and male fertility. J Biol Chem, 301, 108538.